Title
Date

For further details log on website :
http://journals.plos.org/plosone/article?id=10.1371/journal.pone.0144340
Nitrous Oxide (N2O) Emissions by Termites: Does the Feeding Guild Matter?
PublishedDate
- Published: December 10, 2015
- http://dx.doi.org/10.1371/journal.pone.0144340
Abstract
In the tropics, termites are major players in the mineralization of organic matter leading to the production of greenhouse gases including nitrous oxide (N2O). Termites have a wide trophic diversity and their N-metabolism depends on the feeding guild. This study assessed the extent to which N2O emission levels were determined by termite feeding guild and tested the hypothesis that termite species feeding on a diet rich in N emit higher levels of N2O than those feeding on a diet low in N. An in-vitro incubation approach was used to determine the levels of N2O production in 14 termite species belonging to different feeding guilds, collected from a wide range of biomes. Fungus-growing and soil-feeding termites emit N2O. The N2O production levels varied considerably, ranging from 13.14 to 117.62 ng N2O-N d-1 (g dry wt.)-1 for soil-feeding species, with Cubitermes spp. having the highest production levels, and from 39.61 to 65.61 ng N2O-N d-1 (g dry wt.)-1 for fungus-growing species. Wood-feeding termites were net N2O consumers rather than N2O producers with a consumption ranging from 16.09 to 45.22 ng N2O-N d-1 (g dry wt.)-1. Incubating live termites together with their mound increased the levels of N2O production by between 6 and 13 fold for soil-feeders, with the highest increase in Capritermes capricornis, and between 14 and 34 fold for fungus-growers, with the highest increase in Macrotermes muelleri. Ammonia-oxidizing (amoA-AOB and amoA-AOA) and denitrifying (nirK, nirS, nosZ) gene markers were detected in the guts of all termite species studied. No correlation was found between the abundance of these marker genes and the levels of N2O production from different feeding guilds. Overall, these results support the hypothesis that N2O production rates were higher in termites feeding on substrates with higher N content, such as soil and fungi, compared to those feeding on N-poor wood.
Figures
Citation: Brauman A, Majeed MZ, Buatois B, Robert A, Pablo A-L, Miambi E (2015) Nitrous Oxide (N2O) Emissions by Termites: Does the Feeding Guild Matter? PLoS ONE 10(12): e0144340. doi:10.1371/journal.pone.0144340
Editor: John M. Senko, The University of Akron, UNITED STATES
Received: June 3, 2015; Accepted: November 17, 2015; Published: December 10, 2015
Copyright: © 2015 Brauman et al. This is an open access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited
Data Availability: All relevant data are within the paper.
Funding: This study was funded by the project BAMBI (ANR–Métagénome; France).
Competing interests: The authors have declared that no competing interests exist.
Introduction
Nitrous oxide (N2O) is one of the main greenhouse gases in the atmosphere [1] and contributes to the destruction of the ozone layer [2]. Although global annual emissions of N2O have been estimated at 17.7 Tg N [3], the relative contributions from individual sources have not yet been determined.
In tropical terrestrial ecosystems, termites are the most abundant soil macrofauna and have a wide trophic diversity. By virtue of microbial symbionts in their gut, termites play a key role in ecosystem processes such as carbon and nitrogen mineralization [4–6]. Little is known about the extent to which termites contribute to the atmospheric N2O budget in tropical ecosystems, although the metabolic activity of termite gut microbiota maintains steep oxygen gradients within the gut lumen [7], which may help to allow nitrification and denitrification processes to occur simultaneously. These two microbial processes are the principal sources of N2O emissions, either as a by-product of nitrification or as an intermediate in denitrification [8]. Genes that encode for the enzymes involved in the nitrification process include ammonium monooxygenase (amo), hydroxylamine oxidoreductase (hao) and nitrite oxidoreductase (nxr) whereas those that conduct denitrification include nitrite reductases (nirK, nirS), nitric oxide reductase (norB), and nitrous oxide reductase (nosZ). The genes amo, nirK, nirS and nosZ are most commonly used as functional markers to assess the nitrification and denitrification processes [9]. Recently, the use of denitrifying gene abundance as an indicator of greenhouse gas emissions from soils was suggested by Morales et al. [10].
Mineralization of soil nitrogen by termites has mainly been investigated for soil-feeding termite species. Unlike wood-feeding termites which thrive on nutrient-poor material containing as little as 0.05% N [11], soil-feeding species are able to feed on nitrogenous soil components (peptides, proteins and amino acids) as shown by feeding experiments [12–14]. High ammonia concentrations have been detected throughout the termite gut with a considerable accumulation in the posterior hindgut [13]. A study using the 15N-tracer technique showed that denitrification in soil microcosms was stimulated in the presence of termites [15]. Nitrate ingested by soil-feeding termites was denitrified to N2 or reduced to ammonia in the gut [15]. Ngugi and Brune [16] also used the 15N-tracer technique to study the fate of nitrate in the gut compartments of Ophiotermes spp. and two Cubitermes species. Their study was the first report on N2O production in living individuals of soil-feeding termite species and in their gut homogenates after soil-feeding termite mounds had been identified as hot spots of N2O emissions in the African savannah [17]. However, Majeed et al. [18] recently showed that wood-feeding termites were able to take up atmospheric N2O. The fate of nitrogen in termite guts depends on the feeding guild [19]. Therefore, understanding the relationship between feeding behavior and N2O emissions is a critical step in estimating the global contribution of termites to the N2O emission budget in tropical forests.
This study tested the hypothesis that termite species feeding on a diet rich in N emit higher levels of N2O than those feeding on a diet low in N. It set out to assess the extent to which termite feeding behavior affects N2O levels and addressed the differences in N2O production for termite species belonging to various feeding guilds, collected from a wide range of biomes. It also tested for relationships between the feeding guild and the levels of N2O production, the abundance of marker genes for ammonia oxidation (amoA and amoB) and denitrification (nirK, nirS and nosZ) processes, and mineral N concentration in termite guts. So far as we are aware, this is the first study to use a single dataset to compare N2O production rates by termite species belonging to different feeding guilds and to report the presence of ammonia-oxidation and denitrification marker genes in termite guts.
Materials and Methods
Ethics Statement
This study did not involve endangered or protected species. The termites used in this study were collected from the termite rearing laboratory at the Institut de Recherche pour le Développement (IRD) in Bondy (France) with the permission of Dr. Corinne Rouland-Lefèvre, IRD research director–Ile de France.
Termite collection and breeding conditions
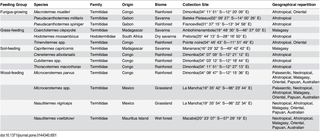
Termites can be classified into four broad feeding guilds: wood-feeders, soil-feeders, fungus-growers and grass-feeders. Fifteen termite species from colonies kept in laboratory conditions belonging to different feeding guilds were used (Table 1). Termite colonies were collected from different sites in tropical and subtropical biomes and kept in the laboratory at the Institute of Research for Development (IRD) at Bondy (France) on a 12-h light/dark cycle. They were kept at 27°C ± 2°C and 80% relative humidity with a constant diet of wood, soil or grass depending on their feeding guild. Worker termites were used for all experiments because they are the most numerous caste and are responsible for all the work in the colony.
Table 1. Termite species studied along with their colony collection sites.
Experimental setup and gas sampling
Experiments were performed using sterile 60 mL serum vials (Wheaton Inc., Millville, USA). The setup comprised (i) live worker termites (0.5 g per vial); (ii) fragments of non-sterilized mound material (5 g) assembled as composite samples from different intact sections of the termite mound to reduce the effect of variations within these biogenic structures, (iii) a combination of live termites and termite non-sterilized mound material and (iv) empty vials.
The vials were sealed at ambient pressure with air-tight sterile butyl rubber septa and aluminum caps and were incubated in the dark at 27°C ± 2°C for 24 h as there is significant mortality for longer incubation times. There were three to five independent replicates for each termite species. Six milliliters of headspace gas were sampled immediately before incubation (T0) using a luer-lock syringe and transferred to 5.9 mL sterilized evacuated Exetainer® vials (Labco Ltd., High Wycombe, England) for further analysis. This was replaced by 6 mL of ambient air to adjust the pressure. After 24-h incubation, the gas was sampled (T24) as described above and kept for analysis.
N2O measurements and up-scaling
The N2O was measured using a gas chromatograph (CP-3800 VARIAN® STAR; Agilent Technologies, Santa Clara, CA, USA), equipped with a 63Ni (15 mCi) electron capturing detector (ECD), as described by Majeed et al. [18]. The equipment was calibrated before the analyses with ambient air and commercial standards of helium and N2O (N48, Air Liquide, France) to give an accurate measurement of the N2O concentrations from 0 to 880 ppm. The detection limit of the GC system was 0.05 ppm. The chromatograms were processed using STAR_Workstation Version 6.0 (Agilent Technologies, Santa Clara, CA, USA) to determine the concentration of N2O in the sample. The results were scaled-up to estimate the total N2O production by termites in tropical forests by multiplying the gas emission rates by an estimate of the termite biomass density as described by Sanderson [20]. In this study, calculations were based on a termite biomass density of 337 kg ha-2 [20].
Determining the ammonium and nitrate concentrations in the termite guts
About 100 mg of whole termite guts were crushed and homogenized in 5 mL of sterilized ultra-pure water. The samples were processed including shaking and centrifugation as described in ISO 14256–2:2005. The ammonium and nitrate in the supernatants of samples were quantified using a San++ Automated Wet Chemistry Analyzer (Skalar Analytical B.V., Netherlands). The ammonium and nitrate concentrations were determined by comparison with the standard curves obtained from samples of known concentrations and ranged from 0 to 4 mg L-1 for ammonium and 0 to 1 mg L-1 for nitrate. Three independent replicates were made for each treatment. Ammonium and nitrate contents were not determined for Crenetermes albotarsalisand Nasutitermes nigriceps.
DNA extraction
Total genomic DNA was extracted from 200 mg whole termite-gut samples. Fast DNA® SPIN Kit for Soil (MP Biomedical, Santa Ana, CA, USA) was used according to the manufacturer’s instructions. Extractions were made in triplicate for each treatment. The extracted DNA was verified by electrophoresis on a 1.5% (w/v) agarose gel which was then stained in a solution of ethidium bromide (0.5 mg L-1) for 15 min, rinsed for 20 min in milli-Q water and checked for the presence of DNA using an UV-transillumination system. The DNA was further quantified by PicoGreen® (Molecular Probes, Inc., Eugene, Oregon, USA). The extracted DNA was stored at -20°C for further analysis.
Quantitative PCR assay
The abundance of 16S rRNA gene and of the key functional marker genes involved in N cycling was estimated by quantitative PCR (qPCR), using a C1000TM thermal cycler (CFX96®, BioRad, Hercules, California) as described by Majeed et al. [21]. The functional marker genes targeted included amoA encoding ammonia monooxygenase for ammonia-oxidizing archaea (AOA) and ammonia-oxidizing bacteria (AOB), and the nirK, nirS and nosZ denitrification genes, encoding copper-containing nitrite reductase, cytochrome cd1 nitrite reductase and nitrous oxide reductase, respectively. Primers targeting the 16S rRNA gene and the functional marker genes and thermal cycling parameters are given in S1 Table. The quantity of DNA was normalized before analysis and tested for PCR inhibition as described by Chapuis-Lardy et al. [22]. Standard curves were obtained from the serial dilutions of a known quantity of linearized pGEM®-T plasmids (Promega, Madison, WI, USA), carrying the targeted DNA sequences. The specificity of the amplified products was confirmed by melting curve analysis. The abundance of 16S rRNA gene and of the functional marker genes was not determined for Thoracotermes macrothorax and Nasutitermes nigriceps.
Statistical analyses
Statistical analyses of N2O emissions by termites were performed using Statistica V 7.1 (StatSoft® Inc., Tulsa, USA). Differences in levels of N2Oemission between termite species were based on the means of three or five replicates for each termite species. Measurements of the abundance of the genes targeted and the mineral N concentration in termite guts were based on eight or nine replicates. Prior to statistical comparison, the data were checked for normal distribution using Shapiro-Wilk test. The means were compared by One-Way Analysis of Variance (ANOVA) and then tested using Tukey’s HSD test with a level of significance of 0.05. The Pearson correlation coefficient was calculated to determine the degree of correlation between the N2O emissions and the abundance of the genes targeted and the mineral N concentration in the termite guts using R version 3.0.0. Results with a value of p< 0.05 were considered significant.
Results
Ammonium and nitrate concentrations in termite guts
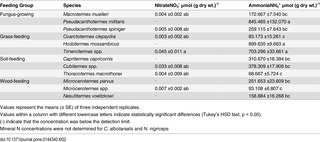
Neither the ammonium nor the nitrate levels were related to termite feeding (Table 2). Ammonia concentrations ranged from 68.7 ± 5.7 to 899.6 ± 9.7 μmol (g dry wt.)-1 in the guts of the termite species studied and were higher than the nitrate concentrations. The highest ammonium concentrations were in the guts of a grass-feeding species, Hodotermes mossambicus, and a fungus-growing species, Pseudacanthotermes militaris. In each guild the concentrations of nitrate in the gut were below detection limit.
Table 2. Mineral N concentrations in gut of different termite species.
Abundance of marker genes for ammonia-oxidizing and denitrifying processes
Functional marker genes for ammonia oxidation (amoA-AOA and amoA-AOB) and denitrification (nirK, nirS and nosZ) processes were successfully amplified from DNA extracts in all the termite guts tested and their copy numbers accounted for less than 10% of the total 16S rRNA gene (except for Microcerotermes parvus). There was no correlation between the abundance of these marker genes and the termite feeding guild to which the termite species belonged (Table 3). With the exception of Crenetermes albotarsalis and M.parvus, the copy numbers of the amoA-AOA marker gene were lower than the abundance of the amoA-AOB marker gene. For the nitrite reductase genes, the abundance of the nirK marker gene was higher than those of the nirS marker gene for all the species tested. The abundance of the nosZmarker gene, coding nitrous oxide reductase, was lower than the nitrite reductase marker genes (nirK and nirS) as indicated by the ratio nosZ/nirK+nirS. Overall, no relationship was found between the gene copy numbers of any of the functional genes tested and the feeding guild to which each species belonged.
Table 3. Copy numbers of 16S rRNA gene and functional marker genes for ammonia-oxidizing and denitrifying processes in gut of different termite species.
Levels of N2O production in different termite feeding guilds
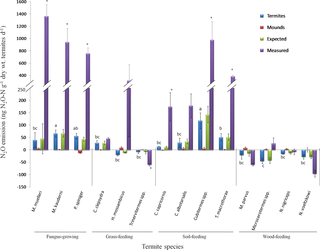
There was no termite mortality in any of the samples at the end of 24-h incubation. Differences in the mean N2O production between termite feeding groups (Fig 1) were statistically significant (P < 0.001). Fungus-growing and soil-feeding termites emit N2O. There were considerable differences in the levels of N2O production, ranging from 13.14 to 117.6 ng N2O-N d-1 (g dry wt.)-1 for soil-feeding species, with Cubitermes spp. having the highest production levels, and from 39.61 to 65.61 ng N2O-N d-1 (g dry wt.)-1 for fungus-growing species. Wood-feeding termites were net N2O consumers rather than N2O producers with a consumption ranging from 16.09 to 45.22 ng N2O-N d-1 (g dry wt.)-1.
Fig 1. Levels of N2O production by termite species belonging to different feeding guilds.
Bars represent means ± SE of three replicates. Expected: indicate N2O production based on the level of emissions measured separately for live termites and for termite mound. Measured: indicate the level of N2O production measured when termites were incubated together with their mound. Different letters mean that there was a statistically significant difference between termite species. * indicate statistically significant differences between level of N2O produced by termites alone and by termite incubated together with mound (Tukey’s HSD test, p < 0.05).
This trend was also observed for two grass-feeding termite species (H. mossambicus and Trinervitermes spp.) but not for Coarctotermes clepsydra which is also a grass-feeder. Little N2O was produced from termite mounds incubated in the absence of live termites. When live termites were incubated with their mounds, N2O production increased by between 7and 13 fold for soil-feeders with the highest increase for Cubitermes spp. and between 14 and 34 fold for fungus-growers with highest increase for M. muelleri (Fig 1). This increase is higher than the sum of N2O produced separately by live termites and by termite mound material (expected production). The differences were statistically significant for fungus-growing termites and for soil-feeding termites except for C. albotarsalis. For the wood-feeding species, M. parvus and Nasutitermes voeltzkowi, an increase of about 3-fold in the level of N2O consumption was obtained in the presence of termite mound material. However, this was not observed for N. nigriceps and Microcerotermes spp. Overall, the levels of N2O production were not correlated with the termite gut mineral N concentration or with the copy numbers of ammonia-oxidizing and denitrifying marker genes in the gut.
Discussion
Are the levels of N2O production affected by the termite feeding guild?
The pattern of N2O production shows that the levels of N2O emissions depend on the termite feeding guild. Termites feeding on soil and fungi produced N2O. In general, while termites feeding on soil and fungi were a source of N2O, wood-feeding and grass-feeding termites were a sink for N2O. Further studies using more species are needed to assess the consumption of N2O in wood-feeding and grass-feeding termites. Overall, our results are consistent with the initial hypothesis that the termite feeding guild is a determinant of N2O production although the average emission levels presented here may not accurately reflect the initial rates at the start of incubation. Cumulative gas production was measured after 24-h incubation in vials (rather than measuring a time series) to ensure that the levels of N2O emissions were above the analytical detection limits. This approach may have underestimated the actual rates. However, the levels of N2O production were not limited by the experimental design and the duration of the experiment was less than the time taken for food to pass through the gut of soil-feeding termites. For the wood-feeding termite species studied, the levels of N2O remaining in the vial headspace after the incubation period accounted for between 10–30% of the total initial quantities. This indicates that, for these termite species and their gut microorganisms, the low N2O concentration in the ambient air did not affect N2O uptake rates through substrate limitation. The zero mortality of termites in all treatments might indicate that not all the oxygen in the vial headspace was depleted by termite respiration.
These results confirm previous findings on N2O production by soil-feeders and N2O consumption by wood-feeders [16, 18]. They also show that some fungus-growing termite species are also a source of N2O and some grass-feeding species may consume atmospheric N2Oin the same way as wood-feeders. The average N2O production level for Cubitermes spp., which is considered to be a true soil-feeding termite, was much higher than for the other termites studied but was about 4-fold lower than reported for Cubitermes ugandensis [16].
There was no correlation between the N2O emission rates and the concentrations of ammonia and nitrate measured in the termite guts. This confirms that pool sizes of metabolites are not good indicators of the emission rate because the mineral-N pool does not adequately reflect N availability [23, 24].
To simulate the termite N2O metabolism under field conditions, live termites were incubated in the presence of their mound material. The higher increase in levels of N2O production under these conditions might be explained by the use of mound material as a food by termites during incubation. Under these conditions, the production of N2O might have remained constant whereas N2O production may have decreased during incubation without mound material as a result of starvation. The increases in N2O production in this study are greater than the increases reported previously in soil microcosms for C. ugandensis and Ophiotermes spp. [16]. This discrepancy might be explained by the differences in the quantities of mineral N ingested by the termites. Mounds of soil-feeding termites have much higher concentrations of mineral N (ammonia and nitrate) than soils [13, 25]. Incubating termites with termite mound fragments results in continuous ingestion of considerable amounts of mineral N and the transformation of the ingested nitrate may increase N2O emissions. It has been established that ingested nitrate is denitrified to N2 or reduced to ammonia in the guts of soil-feeding termites [15, 16]. This study did not measure N2 emissions as it did not set out to determine denitrification pathways in termite guts. Further studies are needed to assess whether or not the denitrification process is complete for various termite species belonging to a particular feeding guild.
Are mineral N and the abundance of marker genes for ammonia-oxidizing and denitrifying bacteria in the guts related to termite diet?
There is little information on mineral-N in the guts of termites belonging to different feeding guilds. However, it has been well documented that there are high ammonia concentrations in the posterior hindgut of soil-feeding species [13, 15, 16]. Our results suggest that high ammonia concentrations in the termite gut are not exclusive to soil-feeding termites as the guts of species belonging to other feeding guilds, such as wood-feeding, grass-feeding and fungus-growing termites, also had high ammonia concentrations. Nitrate concentrations in Cubitermes species were considerably lower than the ammonia concentrations as reported elsewhere [15, 16]. This supports the prevalence of nitrate ammonification processes in the gut of soil-feeding termites [15, 16]. In this study, the ammonia concentrations in the gut of soil-feeding termite species were in the same range as the overall concentrations in the whole gut of Cubitermes species [15]. Higher ammonia concentrations found in soil-feeding and fungus-growing termite species have been explained by the fact that (i) a large proportion of the food of soil-feeding termites is derived from peptides and other nitrogenous humus components of soils [26], (ii) the existence of a nitrate ammonification process in the termite gut [15, 16], (iii) the acquisition of nitrogen by fungus-growing termites from their symbionts, the Termitomyces, a rich source of nitrogen [27].
Given the differences in the levels of N2O production between the termites species studied, the relationship between the feeding guild and the copy numbers of marker genes of ammonia-oxidizing (amoA-AOB; amoA-AOA) and denitrifying bacteria (nirK, nirS and nosZ) in termite guts was investigated. Unlike nifH genes involved in the fixation of atmospheric N2 [28–31], very little is known about marker genes for ammonia-oxidizing and denitrifying processes in termite guts. This study records, for the first time, the presence of the key functional genes involved in these processes in the guts of termite species belonging to a number of different feeding guilds. The abundance of these marker genes did not depend on the feeding guild. This was unexpected since diet can be a major determinant of the bacterial community structure in termite guts [19]. Measuring the abundance of marker genes of ammonia-oxidizers and denitrifiers in the intestinal gut as a whole (as in this study) does not take account of the differences in the composition of the microbial community in each gut compartment which depends on the prevailing physical and chemical conditions [32]. Further studies are needed to determine the expression levels of the marker genes for ammonia-oxidizing and denitrifying bacteria in the various gut compartments. Particular attention should be paid to the genes involved in the dissimilatory nitrate reduction to ammonium since this is the prevailing pathway by which N2O is also produced in termite guts of soil-feeding species [16].
N2O emissions by termites and ecological outcomes
Termites have a high biomass in many tropical biomes and have been recognized as ecosystem engineers mediating various biological soil functions including biogeochemical processes [5]. There is still little information on the contribution of termites to the N2O budget in tropical forests. Soil-feeding termites are more abundant and have greater biomass in tropical forests and humid African forests, whereas Macrotermitinae (fungus-growing termites) are dominant in arid and semi-arid savannas [33]. Taking the highest average levels of N2O production (117.6 ng N-N2O d-1 (g termite dry wt.)-1) recorded in this study for the Cubitermesspp., which is a true soil-feeding termite species, and an estimate of the biomass of soil-feeding termite species in tropical forests (337 kg ha-1; [20]), the production of N2O by termites belonging to the soil-feeding guild was estimated at 0.39 kg N-N2O ha-1 y-1.
This is about 4-fold lower than calculations based on the levels of N2O reported previously for C. ugandensis [16]. However, the contribution of termites to the atmospheric N2O budget is probably higher since the presence of mound material increases N2O production, as shown in this study. A previous study identified the mounds of soil-feeding termites as hot spots of N2O emission in the African savannah [17]. This is confirmed by the positive interaction between termite mound material and live termites for N2O production observed in this study. These findings suggest that scaling up calculations for termite-mediated N2O emissions to the atmosphere should take account of the field conditions.
The N2O uptake by wood-feeding termites is interesting from an ecological point of view. Unfortunately, it was difficult to scale up calculations of N2O consumption by wood-feeding termites as there is a lack of data on their abundance and biomass in the different biomes. It has been established that species in the wood-feeding guild tend to replace soil-feeding termites in response to forest disturbances [4, 34]. Therefore, wood-feeding termites might be important players in scenarios for mitigating N2O emissions into the atmosphere.
The most notable conclusion from this study suggests that the composition of termite feeding guild assemblages in ecosystems should be taken into account when scaling up the contribution of termites to the global budget of N2O emissions. Land-use change might lead to the propagation of one termite feeding guild at the expense of another, and could lead to increased or reduced N2O emissions.
Supporting Information

S1 Table. Primers used in this study for real-time PCR quantification of 16S rRNA genes and functional marker genes for ammonia-oxidizing and denitrification in gut of different termite species.
doi:10.1371/journal.pone.0144340.s001
(DOCX)
Acknowledgments
The authors are grateful to Mr. Laurent Philippot (UMR-SME, Dijon, France) for providing denitrifier primers for qPCR.
Author Contributions
Conceived and designed the experiments: AB MZM EM. Performed the experiments: MZM ALP. Analyzed the data: MZM AB EM. Contributed reagents/materials/analysis tools: AR BB ALP. Wrote the paper: MZM AB EM.
References
- 1.Wuebbles DJ. Nitrous oxide: No laughing matter. Science. 2009; 326: 56–7. doi: 10.1126/science.1179571. pmid:19797649
- 2.Ravishankara AR, Daniel JS, Portmann RW. Nitrous oxide (N2O): the dominant ozone-depleting substance emitted in the 21st century. Science. 2009; 326: 123–5. doi: 10.1126/science.1176985. pmid:19713491
- 3.IPCC. Intergovernmental Panel on Climate Change. IPCC Fourth Assessment Report Climate Change 2007, AR4 Cambridge University Press, Cambridge 2007.
- 4.Bignell DE, Eggleton P. Termites in ecosystems. In: Abe T, Bignell DE, Higashi H, editors. Termites: Evolution, Sociality, Symbiosis, Ecology. Kluwer, Dordrecht. 2000. pp. 363–408.
- 5.Bignell DE. Termites as soil engineers and soil processors. In: König H, Varma A, editors. Intestinal microorganisms of termites and other invertebrates: Springer, Berlin Heidelberg. 2006. pp. 183–220.
- 6.Jouquet P, Traoré S, Choosai C, Hartmann C, Bignell D. Influence of termites on ecosystem functioning. Ecosystem services provided by termites. Eur J Soil Biol. 2011; 47: 215–22. doi: 10.1016/j.ejsobi.2011.05.005
- 7.Brune A, Emerson D, Breznak JA. The termite gut microflora as an oxygen sink–microelectrode determination of oxygen and pH gradients in guts of lower and higher termites. Appl Environ Microbiol. 1995; 61: 2681–7. pmid:16535076
- 8.Conrad R. Soil microorganisms as controllers of atmospheric trace gases (H2, CO, CH4, OCs, N2O, and NO). Microbiol Rev. 1996; 60: 609–40. pmid:8987358
- 9.Levy-Booth DJ, Prescott CE, Grayston SJ. Microbial functional genes involved in nitrogen fixation, nitrification and denitrification in forest ecosystems. Soil Biol Biochem. 2014; 75: 11–25. doi: 10.1016/j.soilbio.2014.03.021
- 10.Morales SE, Cosart T, Holben WE. Bacterial gene abundances as indicators of greenhouse gas emission in soils. Isme J. 2010; 4: 799–808. doi: 10.1038/ismej.2010.8. pmid:20182521
- 11.Tayasu I, Sugimoto A, Wada E, Abe T. Xylophagous termites depending on atmospheric nitrogen. Naturwissenschaften. 1994; 81: 229–31. doi: 10.1007/s001140050063
- 12.Ji R, Kappler A, Brune A. Transformation and mineralization of synthetic 14C-labeled humic model compounds by soil-feeding termites. Soil Biol Biochem. 2000; 32: 1281–91. doi: 10.1016/s0038-0717(00)00046-8
- 13.Ji R, Brune A. Nitrogen mineralization, ammonia accumulation, and emission of gaseous NH3 by soil-feeding termites. Biogeochemistry. 2006; 78: 267–83. doi: 10.1007/s10533-005-4279-z
- 14.Ji R, Brune A. Transformation and mineralization of 14C-labeled cellulose, peptidoglycan, and protein by the soil-feeding termite Cubitermes orthognathus. Biol Ferti Soil. 2001; 33: 166–74. doi: 10.1007/s003740000310
- 15.Ngugi DK, Ji R, Brune A. Nitrogen mineralization, denitrification, and nitrate ammonification by soil-feeding termites: a 15N-based approach. Biogeochemistry. 2011; 103: 355–69. doi: 10.1007/s10533-010-9478-6
- 16.Ngugi DK, Brune A. Nitrate reduction, nitrous oxide formation, and anaerobic ammonia oxidation to nitrite in the gut of soil-feeding termites (Cubitermes and Ophiotermesspp.). Environ Microbiol. 2012; 14: 860–71. doi: 10.1111/j.1462-2920.2011.02648.x. pmid:22118414
- 17.Brümmer C, Papen H, Wassmann R, Bruggemann N. Termite mounds as hot spots of nitrous oxide emissions in South-Sudanian savanna of Burkina Faso (West Africa). Geophys Res Lett. 2009; 36. doi: 10.1029/2009gl037351
- 18.Majeed MZ, Miambi E, Robert A, Bernoux M, Brauman A. Xylophagous termites: A potential sink for atmospheric nitrous oxide. Eur J Soil Biol. 2012; 53:121–5. doi: 10.1016/j.ejsobi.2012.10.002
- 19.Brune A. Symbiotic digestion of lignocellulose in termite guts. Nat Rev Microbiol. 2014; 12: 168–80. doi: 10.1038/nrmicro3182. pmid:24487819
- 20.Sanderson MG. Biomass of termites and their emissions of methane and carbon dioxide: A global database. Glob Biogeochem Cy. 1996; 10: 543–57. doi: 10.1029/96gb01893
- 21.Majeed MZ, Miambi E, Barois I, Blanchart E, Brauman A. Emissions of nitrous oxide from casts of tropical earthworms belonging to different ecological categories. Pedobiologia. 2012; 56: 49–58. doi: 10.1016/j.pedobi.2012.10.003
- 22.Chapuis-Lardy L, Brauman A, Bernard L, Pablo AL, Toucet J, Mano MJ, et al. Effect of the endogeic earthworm Pontoscolex corethrurus on the microbial structure and activity related to CO2 and N2O fluxes from a tropical soil (Madagascar). Appl Soil Ecol. 2010; 45: 201–8. doi: 10.1016/j.apsoil.2010.04.006
- 23.Attard E, Recous S, Chabbi A, De Berranger C, Guillaumaud N, Labreuche J, Philippot L, Schmid B, Le Roux X. Soil environmental conditions rather than denitrifier abundance and diversity drive potential denitrification after changes in land uses, Global Chang Biol. 2011; 17: 1975–1989. doi: 10.1111/j.1365-2486.2010.02340.x
- 24.Bru D, Ramette A, Saby NPA, Dequiedt S, Ranjard L, Jolivet C, et al. Determinants of the distribution of nitrogen-cycling microbial communities at the landscape scale. Isme J. 2011; 5: 532–42. doi: 10.1038/ismej.2010.130. pmid:20703315
- 25.Ndiaye D, Lensi R, Lepage M, Brauman A. The effect of the soil-feeding termite Cubitermes niokoloensis on soil microbial activity in a semi-arid savanna in West Africa. Plant and Soil. 2004; 259: 277–86. doi: 10.1023/b:plso.0000020980.50095.e1
- 26.Kappler A, Ji R, Brune A. Synthesis and characterization of specifically 14C-labeled humic model compounds for feeding trials with soil-feeding termites. Soil Biol Biochem. 2000; 32: 1271–80. doi: 10.1016/s0038-0717(00)00047-x
- 27.Rouland-Lefèvre C. Symbiosis with fungi. In: Abe T, Bignell DE, Higashi M, editors. Termites: evolution, sociality, symbioses, ecology, Kluwer Academic Publishers, Dordrecht. 2000. pp. 289–306.
- 28.Ohkuma M, Noda S, Kudo T. Phylogenetic diversity of nitrogen fixation genes in the symbiotic microbial community in the gut of diverse termites. Appl Environ Microbiol. 1999; 65: 4926–34. pmid:10543805
- 29.Ohkuma M, Noda S, Usami R, Horikoshi K, Kudo T. Diversity of nitrogen fixation genes in the symbiotic intestinal microflora of the termite Reticulitermes speratus. Appl Environ Microbiol. 1996; 62: 2747–52. pmid:16535372
- 30.Yamada A, Inoue T, Noda S, Hongoh Y, Ohkuma M. Evolutionary trend of phylogenetic diversity of nitrogen fixation genes in the gut community of wood-feeding termites. Mol Ecol. 2007; 16: 3768–77. pmid:17850544 doi: 10.1111/j.1365-294x.2007.03326.x
- 31.Desai MS, Brune A. Bacteroidales ectosymbionts of gut flagellates shape the nitrogen-fixing community in dry-wood termites. ISME J. 2012; 6: 1302–13. doi: 10.1038/ismej.2011.194. pmid:22189498
- 32.Köhler T, Dietrich C, Scheffrahn RH, Brune A. High-resolution analysis of gut environment and bacterial microbiota reveals functional compartmentation of the gut in wood-feeding higher termites (Nasutitermes spp.). Appl Environ Microbiol. 2012; 78: 4691–4701. doi: 10.1128/AEM.00683-12. pmid:22544239
- 33.Wood T, Sands W. Role of termites in ecosystems, In: Brian MV, editor. Production Ecology of Ants and Termites, International Biological Programme, Cambridge University Press, Cambridge. 1978. pp. 245–292.
- 34.Eggleton P, Bignell DE, Sands WA, Mawdsley NA, Lawton , Wood TG, et al. The diversity, abundance and biomass of termites under differing levels of disturbance in the Mbalmayo Forest Reserve, Southern Cameroon. Philos T Roy Soc B. 1996; 351: 51–68. doi: 10.1098/rstb.1996.0004
For further details log on website :
http://journals.plos.org/plosone/article?id=10.1371/journal.pone.0144340









No comments:
Post a Comment