Published Date
How to cite this article:
J.C.F. Bong, P.J.H. King, K.H. Ong and N.M. Mahadi, 2012. Termites Assemblages in Oil Palm Plantation in Sarawak, Malaysia. Journal of Entomology, 9: 68-78.
DOI: 10.3923/je.2012.68.78
URL: http://scialert.net/abstract/?doi=je.2012.68.78
Author
J.C.F. Bong, P.J.H. King, K.H. Ong and N.M. Mahadi
INTRODUCTION
Termites are one of the popular studied insect to the research community nowadays. This is mainly due to the economic damage caused by their voracious appetite for wood as well as their potential in biomass conversion to fuel. Termites are known as gold mine of genes, microbes and enzymes that are potentially useful in converting biomass into ethanol and methanol that are highly value as alternative fuel to petroleum. Thus, intensive metagenomic and transcriptomic research are carried out worldwide on termites (Warnecke et al., 2007; Tartar et al., 2009).
There are about 2650 species of termites that have so far been documented worldwide (Kambhampati and Eggleton, 2000). Great diversity in termites provides not only huge among of genes and alleles as well as balance in ecosystem equilibrium (Turner, 2004) and enhancement in ecosystem production (Pringle et al., 2010). Different species have their own uniqueness in diet preference, habitat, micro niche and species specific intestinal microbes hence disparately affect the ecosystem carbon flow and productivity. The study in African’s savanna found that the Odontotermes termites help to improve soil fertility, elevated level of phosphorus and nitrogen as well as greatly enhanced plants and animals’ activity at local level (Pringle et al., 2010). Termites are known to lead a very cryptic life in the soil or wood, building tunnels, aggregates soil into mounds and transferring particles from different soil layers. Such behaviors have great economic impact to human as it alters the physical structure of agricultural lands and wood structures. Termites create and modify microhabitats for smaller terrestrial organism and change soil physical property which includes soil water holding capacity, aeration, drainage and stability. This bioturbation is important as compacted and encrusted soils have low water absorptivity and hence not efficient in supporting plant life. A lot of researches have acknowledged that termites as soil ecosystem engineers (Stork and Eggleton, 1992; Jones et al., 1994; Lawton, 1996; Lavelle et al., 1997). They also form faeces which are organomineral complexes, stable over periods of months or more (Lavelle et al., 1997).
Majority of termites are fed on semi degraded wood material and few are known to feed on living trees, leaf litter, soil, lichens and animal faeces. Based on their feeding behavious, termites can be divided into five different groups (Donovan et al., 2001). The first group of termites are humus and mineral soil feeder; Group 2 obtain their nutrient and energy from wood (fresh or rotten); Third group prefers soil humus comprise of litter-soil mixture, extremely decayed wood, or soil adjacent to logs; Group 4 feed predominantly on leaf litter and small items of woody trash; and Group 5 feed on lichens and other free-living non-vascular plants which graze from the surface of tree trunks, example: Hospitalitermes. Due to the different feeding habit, each termite species plays significant different role and strive at different efficient level in their ecosystem. Depends on the biotic and abiotic elements present in the ecosystem, the abundance of each termite species should be different at different ecosystem. A large area of oil plantations in Sarawak were established on former peat swamp area (estimated 20% of total planted area). Majority of these areas were reported infested by termites. Hence, this study aimed to determine the termite diversity in Sarawak oil palm plantation that was established on peat.
MATERIALS AND METHODS
Site description and sampling technique: The termite diversity study was carried out at Sunda region in Semanok oil palm estate, located in Tatau, Bintulu, Sarawak, Malaysia (Latitude: 3°01’00.20”N and Longtitute: 112°52’48.03”E) which was previously a peat area with depth greater than 1.5 m and soil acidity at around 4-5 pH. The study was carried out for seven months started in early August 2009 and ended in February 2010. Standardized sampling protocol was applied that is proposed by Jones and Eggleton (2000). This method is rapid and cost-effective for termite assemblage structure study.
A transect sampling was conducted in this study. This transect was 100 m long and 6 m wide with 20 contiguous sections. Each section was sampled sequentially. Every microsites including soil surface material, such as litter, humus, dead logs, tree stump, branches, twigs, rotten logs, subterranean nest, mounds, carton nest, gallery on vegetation or rock and arboreal nests up to a height of 2 m above ground level, were carefully investigated in detail and all in sight termites were captured. Termites were collected with insect aspirators and soft brush. Termites were then preserved under -80°C or 70% alcohol. Termite identification was carried out based on termites’ external morphology primarily of the soldier caste. This was carefully done under the aid of dissecting microscope.
Termite identification: All termite colonies collected were coded and carefully divided into groups based on Prestwich (1984) classification (Table 1). External features of a termite soldier were measured and compared with records of Thapa (1981).
A total of 15 morphometric characters were used (Table 2) to identify the specimen collected. Ten individuals (soldier) for each termite colony were measured. All the measurements were recorded in millimeters.
Data analysis: All data were subjected to Analysis of Variance (ANOVA) using SAS version 9.0. The differences between mean of morphological measurement among the same genera was compared using Tukey test at p>0.05.
RESULTS AND DISCUSSION
A total of 13 species of termites were identified. Species T1 (Fig. 1a) which was collected from an infested oil palm fruit bunch and identified as wood feeder and wood nesting termites. T1 soldiers have brownish yellow oval-shaped head capsule with scattered bristles. They had long saber-shaped Slashing Mandible (SM) and were strongly incurved at apical. Their left mandibles have three crenulations at base while right mandibles were without crenulations. Labrums were yellow in color and had a distinct pair of long bristles on the tip and a few bristles on the disc. T1 soldiers also have large fontanelle with two long bristles at the rim (Fig. 2). Their antennas were with 16 segments and abdomens had ten segments. Soldiers T1 would excrete a white fluid from their fontanelle when being disturbed or attacked.
The morphological characters of T1 suggested T1 as Coptotermes curvignathus (Holmgren). This species is recognized as a pest to agricultural and forestry tree species, causing economical damage to both young and mature trees (Tho and Kirton, 1992; Tho, 1992; Kirton et al., 1999).
Species coded as T2 (Fig. 1b) was collected from rotten wood in the study area. T2 shared high degree of similarity with T1. However, differ from T1, soldier of T2 has antenna with 14 segments instead of 16 and T1 soldier was significantly larger (p>0.05, data not shown) in size as compare to T2, with head length to side base of mandible at 1.64±0.014 mm for T1 and 1.21±0.010 mm for T2; head width at 1.48±0.019 mm for T1 and 1.06±0.008 mm for T2 and head length for T1 at 0.95±0.022 mm and T2 at 0.73±0.009 mm. T2 was identified as Coptotermes sepangensis Krishna.
Colony T3 (Fig. 1c) was found under rotten wood, partially decomposed materials and below the ground with lichens growing on the surface. T3 was recognized as wood feeder and wood nesting termites. The head capsules of T3 soldier were dorsally flat and broad. Their mandibles were the crushing mandibular type and were short and strongly bent inwards in the middle with broad bases. Left mandible had a long finger-like apical tooth and two marginal teeth while right mandible had only one apical tooth and one marginal tooth. The fontanelle of T3 soldiers was minute and circular. Their antennas had 13 segments and abdomens were ten segmented. T3 was morphometrically identified as Parrhinotermes aequalis (Havilandi) and is the only Parrhinotermes species found in the study site.
Colonies T4, T5, T6 and T7 built their galleries in the rotten wood and damp wood on the surface and under the peat. Based on their feeding habit, they were all wood feeders and wood-nesting builders. They all exhibit soldier dimorphism (Fig. 3) and have crushing mandibles. The general characteristics of T4 (Fig. 3a), T5 (Fig. 3b), T6 (Fig. 3c) and T7 (Fig. 3d) suggested that they were belonging to Schedorhinotermesgenus. However, these colonies displayed significant different features as shown in Table 3. Soldiers of T4 have significantly (p>0.05) larger head length, head width, head height and length of mandibles as compared to T5 and T6 (Table 3). When compared with T7, soldiers of T4 have significantly (p>0.05) longer head length, head width and left mandible (Table 3). In addition, T4 had 18 segmented antennal while T5 had 16-17 segmented. This characteristic of T4 had not been reported in any Schedorhinotermes species therefore T4 could be a novel termite species. Based on the morphology, T5 was identified as Schedorhinotermes brevialatus (Haviland), T6 as Schedorhinotermes javanicus Kemner and T7 as Schedorhinotermes sarawakensis (Holmgren).
T8 built their galleries under the peat (approximately 10 cm from the surface) and they were also found below the ground with lichens grow on the surface. T8 were also often found sharing the same habitat with Schedorhinotermes species. Based on their habitat and feeding sources, T8 (Fig. 4a) was recognized as a soil feeder and subterranean-nesting termites. T8 soldiers have elongated head capsule with length twice as long as their width. Their head were yellowish brown with a reddish tinge and covered by a few scattered bristles. Mandibles of T8 were classified as asymmetrical snapping, short and strongly asymmetrical. Left mandibles of T8 soldiers were strongly twisted at the middle with rounded tip while their right mandible were slightly shorter than left mandible, blade-like and with the tip slightly sharply pointed and curved outward. T8 soldiers had 14 brown segmented antennals and ten segmented abdomen. T8 were identified as Pericapritermes dolichocephalus (John). T9 (Fig. 4b) showed morphological similarities with T8 but were distinctly different in term of size. T9 soldiers have significantly (p>0.05) larger head length, head width, head height and length of mandibles than T8 (Table 4) and was identified as Pericapritermes latignathus (Holmgren).
Colony T10 (Fig. 5a) was detected based on their mud-trails formed from the ground to the top of the trunks and fronds of oil palm. T10 had their built their nest in oil palm fruit bunches and were found to be an interface feeder. T10 was morphologically classified as glue squirting termites. Head of T10 soldiers (without rostrum) were oval-like and the width is longer wider than length. Their heads were reddish yellow to brownish yellow with a pair of bristles on posterior of the head and four short bristles on side of rostrum base. Dorsal profile of the head capsule was nearly straight.
Rostrum of T10 was short and cone shaped and the tip of the rostrum was reddish brown and darker than the head. Mandibles of T10 were short apical processes without rudimentary tooth. T10 has 13 antennal segments and ten abdominal segments. T10 was identified as Nasutitermes havilandi (Desneux).
T11 (Fig. 5b) was collected from rotten wood found in peat and no mud-trails detected on the trunks and fronds of oil palm. Based on their habitat and feeding sources, T11 was identified as wood feeder and wood-nesting termites. Similar to T10, T11 was morphologically classified as glue squirting termites. Head of T11 (without rostrum) was subcircular. However, differ from T10, dorsal profile of T11 head capsule showed a shallow concave at the middle. Some specimens were weakly indented in the middle at the posterior margin of the head. T11 was identified as Nasutitermes longinasoides Thapa.
Similar to T10 and T11, T12 was morphologically classified as glue squirting termite however, they were different in size (Table 5). T11 soldiers were the largest among the three colonies and the T10 were the smallest.
Soldiers of T11 have significantly (p>0.05) longer head width, head height, head length and rostrum length as compared with T10 and T12 (Table 5). However, T11 has significantly (p>0.05) smaller rostrum-head index than T10 and T12. T12 (Fig. 5c) were identified as Nasutitermes matangensiformis Holmgren.
Colony T13 (Fig. 6) was found in rotten oil palm fronds and rotten wood without any mud trails. T13 also built their galleries under the peat (approximately 10 cm from the surface). Based on their habitat and feeding sources, T13 was recognized as an interface feeder termite. T13 was morphologically classified as a glue squirting termite. T13 soldiers had reddish brown head without hairs. Dorsal profile of T13 head capsule showed a weak depression in the middle. Its rostrum was long, cone-shaped and the size was longer than head capsule. T13 had 14 yellowish brown antennal segments and ten abdominal segments. Based the morphological characters, T13 was identified as Havilanditermes atripennis (Haviland).
Based on the morphometric identification, a total of 13 species of termite from six different genera were found in Sarawak Oil Palm Plantation peat zone (Fig. 7). Genus of Schedorhinotermes were the most diverse in term of species (4 spp. or 31%), followed by Nasutitermes (3 spp. or 23%), Coptotermes (2 spp. or 15%), Pericapritermes (2 spp. or 15%), Havilanditermes (1 sp. or 8%) and Parrhinotermes (1 sp. or 8%).
This study found that species of Coptotermes are one of the most commonly encountered species in oil palm plantation which is established on former peat land. Coptotermes curvignathus was abundantly found in the studied site. This is because the species is known to have voracious appetite for living trees instead of dead or decayed wood. The oil palm plantation provides a generous feeding ground for this species. In contrast, Coptotermes was less dominant in rural area where termite infestation was reported (Kirton, 2005). The other species of Coptotermes, C. gestroi which is responsible for most of the urban dwelling in Malaysia (Kirton and Brown, 2003) was not found in the study areas.
These reaffirm the differences in feeding preferences of these two species. Coptotermes gestroi prefers dead wood and C. curvignathusprefers to feed on living trees.
All termites that were found in the study sites were exclusively from Rhinotermitidae and Termitidae families. Rhinotermitidae was outnumbering species of Termitidae in the study areas. This is in contrast with Eggleton (2000) that studied on termite assemblage in a conserved Dipterocarp forest in Sabah. They reported that the Rhinotermitidae made up only moderate proportion of the whole assemblages in dipterocarp forest and the most dominant termite fauna were the termes-group soil feeders and Nasutitermes. The contrast findings between Eggleton (2000) and ours showed that agricultural practices may have some effect on the termite assemblages especially on the wood feeder termites as well as Termitidae species which are also known as higher termites. In our study, the wood feeders constitute 54% of the total termites found in the studied areas (Fig. 8), outnumbering the soil and soil/wood feeders. These may be due to changes in soil properties as consequences of agricultural practices which often have numerous repercussions on the physical and biological properties on soil. Studies have shown that the species varieties and abundance of soil microbes in tropical agricultural lands is less than 50% of that of the primary forest originally on the land (Matson et al., 1997) and soil carbon can drop more than 50% within five years when tropical soils were converted for agricultural purposes. In addition to these, when tropical soils are cultivated, they become substantially compacted and surface albedo is increased leading to temperature increase and decreases in precipitation. These all in all may have transformed the ecosystem into a hostile environment for certain species of termites especially species of soil termites and higher termites which often depending on soil microbes or fungi.
The result also showed that lesser negative impact of agricultural practices on wood feeding termites as the cultivation provides food to wood feeder termites. Rhinotermitidae species which often feed on economic importance crops (Edwards and Mill, 1986; French et al., 1986; Su and Scheffrahn, 1998) hence was observed to have the biggest biomass density in this studied agrisystem. However, the faunistics difference before the cultivation intervention between this study site and conserved dipterocarp forest should be taken note of as both sites are in Sunda region.
CONCLUSION
The result of this study has shown that the wood feeder termites are the main species in oil palm plantation that is situated on former peat swamp. The major pest is C. curvignathus which infests living agricultural plants. Scavengers such as Schedorhinotermes spp., Pericapritermes spp., H. atripennis, C. sepangensis and P. aequalis were commonly found in oil palm plantation however, they induce no economical damage to oil palm trees and probably may have positively improved soil condition. These findings suggest that chemical termiticides draining which is a common practice in oil palm plantations may be unnecessary. This is because the practice may negatively affect the termites’ assemblages in agricultural land and causes unnecessary ecology drift. Baiting system using rubber wood chunk to attract C. curvignathus will be a better approach and cause less damage to soil and higher termites.
ACKNOWLEDGMENTS
This research was supported by the RUGS grant (Grant No. 91969 to Patricia King Jie Hung) from Universiti Putra Malaysia, IRPA grant (Grant No. 09-02-02-2008-BTK/ER-36 to Nor Muhammad Mahadi) from the Ministry of Science, Technology and Environment, Malaysia and e-Science grant (Grant No. 05-01-04-SF0516 to Joseph Choon Fah Bong) from the Ministry of Science Technology and Innovations, Malaysia. The authors would like to thank Wong Chi Kang for his their technical help.
For further details log on website :
http://scialert.net/fulltext/?doi=je.2012.68.78&org=10
| Received: June 05, 2011; Accepted: October 16, 2011; Published: December 12, 2011 | |
J.C.F. Bong, P.J.H. King, K.H. Ong and N.M. Mahadi, 2012. Termites Assemblages in Oil Palm Plantation in Sarawak, Malaysia. Journal of Entomology, 9: 68-78.
DOI: 10.3923/je.2012.68.78
URL: http://scialert.net/abstract/?doi=je.2012.68.78
Author
J.C.F. Bong, P.J.H. King, K.H. Ong and N.M. Mahadi
INTRODUCTION
Termites are one of the popular studied insect to the research community nowadays. This is mainly due to the economic damage caused by their voracious appetite for wood as well as their potential in biomass conversion to fuel. Termites are known as gold mine of genes, microbes and enzymes that are potentially useful in converting biomass into ethanol and methanol that are highly value as alternative fuel to petroleum. Thus, intensive metagenomic and transcriptomic research are carried out worldwide on termites (Warnecke et al., 2007; Tartar et al., 2009).
There are about 2650 species of termites that have so far been documented worldwide (Kambhampati and Eggleton, 2000). Great diversity in termites provides not only huge among of genes and alleles as well as balance in ecosystem equilibrium (Turner, 2004) and enhancement in ecosystem production (Pringle et al., 2010). Different species have their own uniqueness in diet preference, habitat, micro niche and species specific intestinal microbes hence disparately affect the ecosystem carbon flow and productivity. The study in African’s savanna found that the Odontotermes termites help to improve soil fertility, elevated level of phosphorus and nitrogen as well as greatly enhanced plants and animals’ activity at local level (Pringle et al., 2010). Termites are known to lead a very cryptic life in the soil or wood, building tunnels, aggregates soil into mounds and transferring particles from different soil layers. Such behaviors have great economic impact to human as it alters the physical structure of agricultural lands and wood structures. Termites create and modify microhabitats for smaller terrestrial organism and change soil physical property which includes soil water holding capacity, aeration, drainage and stability. This bioturbation is important as compacted and encrusted soils have low water absorptivity and hence not efficient in supporting plant life. A lot of researches have acknowledged that termites as soil ecosystem engineers (Stork and Eggleton, 1992; Jones et al., 1994; Lawton, 1996; Lavelle et al., 1997). They also form faeces which are organomineral complexes, stable over periods of months or more (Lavelle et al., 1997).
Majority of termites are fed on semi degraded wood material and few are known to feed on living trees, leaf litter, soil, lichens and animal faeces. Based on their feeding behavious, termites can be divided into five different groups (Donovan et al., 2001). The first group of termites are humus and mineral soil feeder; Group 2 obtain their nutrient and energy from wood (fresh or rotten); Third group prefers soil humus comprise of litter-soil mixture, extremely decayed wood, or soil adjacent to logs; Group 4 feed predominantly on leaf litter and small items of woody trash; and Group 5 feed on lichens and other free-living non-vascular plants which graze from the surface of tree trunks, example: Hospitalitermes. Due to the different feeding habit, each termite species plays significant different role and strive at different efficient level in their ecosystem. Depends on the biotic and abiotic elements present in the ecosystem, the abundance of each termite species should be different at different ecosystem. A large area of oil plantations in Sarawak were established on former peat swamp area (estimated 20% of total planted area). Majority of these areas were reported infested by termites. Hence, this study aimed to determine the termite diversity in Sarawak oil palm plantation that was established on peat.
MATERIALS AND METHODS
Site description and sampling technique: The termite diversity study was carried out at Sunda region in Semanok oil palm estate, located in Tatau, Bintulu, Sarawak, Malaysia (Latitude: 3°01’00.20”N and Longtitute: 112°52’48.03”E) which was previously a peat area with depth greater than 1.5 m and soil acidity at around 4-5 pH. The study was carried out for seven months started in early August 2009 and ended in February 2010. Standardized sampling protocol was applied that is proposed by Jones and Eggleton (2000). This method is rapid and cost-effective for termite assemblage structure study.
A transect sampling was conducted in this study. This transect was 100 m long and 6 m wide with 20 contiguous sections. Each section was sampled sequentially. Every microsites including soil surface material, such as litter, humus, dead logs, tree stump, branches, twigs, rotten logs, subterranean nest, mounds, carton nest, gallery on vegetation or rock and arboreal nests up to a height of 2 m above ground level, were carefully investigated in detail and all in sight termites were captured. Termites were collected with insect aspirators and soft brush. Termites were then preserved under -80°C or 70% alcohol. Termite identification was carried out based on termites’ external morphology primarily of the soldier caste. This was carefully done under the aid of dissecting microscope.
Termite identification: All termite colonies collected were coded and carefully divided into groups based on Prestwich (1984) classification (Table 1). External features of a termite soldier were measured and compared with records of Thapa (1981).
| Table 1: | Prestwich classification of termite |
 | |
| Table 2: | External morphomatrix of termite (soldier) |
 | |
Data analysis: All data were subjected to Analysis of Variance (ANOVA) using SAS version 9.0. The differences between mean of morphological measurement among the same genera was compared using Tukey test at p>0.05.
RESULTS AND DISCUSSION
A total of 13 species of termites were identified. Species T1 (Fig. 1a) which was collected from an infested oil palm fruit bunch and identified as wood feeder and wood nesting termites. T1 soldiers have brownish yellow oval-shaped head capsule with scattered bristles. They had long saber-shaped Slashing Mandible (SM) and were strongly incurved at apical. Their left mandibles have three crenulations at base while right mandibles were without crenulations. Labrums were yellow in color and had a distinct pair of long bristles on the tip and a few bristles on the disc. T1 soldiers also have large fontanelle with two long bristles at the rim (Fig. 2). Their antennas were with 16 segments and abdomens had ten segments. Soldiers T1 would excrete a white fluid from their fontanelle when being disturbed or attacked.
 | |
| Fig. 1(a-c): | Soldier (left) and worker (right) of: (a) T1, Coptotermes curvignathus; (b) T2, Coptotermes sepangensis; and T3, Parrhinotermes aequalis |
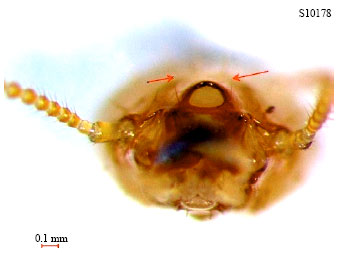 | |
| Fig. 2: | Distinctive fontanelle and one pair of hair near the rim of the fontanelle (pointed by arrows) of Coptotermes curvignathus |
Species coded as T2 (Fig. 1b) was collected from rotten wood in the study area. T2 shared high degree of similarity with T1. However, differ from T1, soldier of T2 has antenna with 14 segments instead of 16 and T1 soldier was significantly larger (p>0.05, data not shown) in size as compare to T2, with head length to side base of mandible at 1.64±0.014 mm for T1 and 1.21±0.010 mm for T2; head width at 1.48±0.019 mm for T1 and 1.06±0.008 mm for T2 and head length for T1 at 0.95±0.022 mm and T2 at 0.73±0.009 mm. T2 was identified as Coptotermes sepangensis Krishna.
 | |
| Fig. 3(a-d): | Soldier (left) and worker (right) of: (a) T4, Schedorhinotermes sp.; (b) T5,Schedorhinotermes brevialatus; (c) T6, Schedorhinotermes javanicus; and (d) T7, Schedorhinotermes sarawakensis |
Colonies T4, T5, T6 and T7 built their galleries in the rotten wood and damp wood on the surface and under the peat. Based on their feeding habit, they were all wood feeders and wood-nesting builders. They all exhibit soldier dimorphism (Fig. 3) and have crushing mandibles. The general characteristics of T4 (Fig. 3a), T5 (Fig. 3b), T6 (Fig. 3c) and T7 (Fig. 3d) suggested that they were belonging to Schedorhinotermesgenus. However, these colonies displayed significant different features as shown in Table 3. Soldiers of T4 have significantly (p>0.05) larger head length, head width, head height and length of mandibles as compared to T5 and T6 (Table 3). When compared with T7, soldiers of T4 have significantly (p>0.05) longer head length, head width and left mandible (Table 3). In addition, T4 had 18 segmented antennal while T5 had 16-17 segmented. This characteristic of T4 had not been reported in any Schedorhinotermes species therefore T4 could be a novel termite species. Based on the morphology, T5 was identified as Schedorhinotermes brevialatus (Haviland), T6 as Schedorhinotermes javanicus Kemner and T7 as Schedorhinotermes sarawakensis (Holmgren).
 | |
| Fig. 4(a-b): | Soldier (left) and worker (right) of: (a) T8, Pericapritermes dolichocephalus; and (b) T9, Pericapritermes latignathus |
| Table 3: | Characteristics comparison (mm) between ten minor soldiers of T4, T5, T6 and T7 |
 | |
| Values are as Mean±SD. Means with the same letter across row are not significantly different p>0.05; LHM, Length of head to side base of mandible; MWP: Maximum width of head; HOH: Height of head; LLM: Length of left mandible; LRM: Length of right mandible | |
Colony T10 (Fig. 5a) was detected based on their mud-trails formed from the ground to the top of the trunks and fronds of oil palm. T10 had their built their nest in oil palm fruit bunches and were found to be an interface feeder. T10 was morphologically classified as glue squirting termites. Head of T10 soldiers (without rostrum) were oval-like and the width is longer wider than length. Their heads were reddish yellow to brownish yellow with a pair of bristles on posterior of the head and four short bristles on side of rostrum base. Dorsal profile of the head capsule was nearly straight.
 | |
| Fig. 5(a-c): | Soldier (left) and worker (right) of: (a) T10, Nasutitermes havilandi; (b) T11, Nasutitermes longinasoides; and T12, Nasutitermes matangensiformis |
| Table 4: | Characteristics comparison (mm) between ten soldiers of T8 and T9 |
 | |
| Values are as Mean±SD. Means with the same letter across row are not significantly different p>0.05; *Sample codes correspond to those in Table 3 | |
T11 (Fig. 5b) was collected from rotten wood found in peat and no mud-trails detected on the trunks and fronds of oil palm. Based on their habitat and feeding sources, T11 was identified as wood feeder and wood-nesting termites. Similar to T10, T11 was morphologically classified as glue squirting termites. Head of T11 (without rostrum) was subcircular. However, differ from T10, dorsal profile of T11 head capsule showed a shallow concave at the middle. Some specimens were weakly indented in the middle at the posterior margin of the head. T11 was identified as Nasutitermes longinasoides Thapa.
Similar to T10 and T11, T12 was morphologically classified as glue squirting termite however, they were different in size (Table 5). T11 soldiers were the largest among the three colonies and the T10 were the smallest.
 | |
| Fig. 6: | Soldier (left) and worker (right) of T13, Havilanditermes atripennis |
| Table 5: | Characteristics comparison (mm) between ten soldiers of T10, T11 and T12 |
 | |
| Values are as Mean±SD. Means with the same letter across row are not significantly different p>0.05 | |
Colony T13 (Fig. 6) was found in rotten oil palm fronds and rotten wood without any mud trails. T13 also built their galleries under the peat (approximately 10 cm from the surface). Based on their habitat and feeding sources, T13 was recognized as an interface feeder termite. T13 was morphologically classified as a glue squirting termite. T13 soldiers had reddish brown head without hairs. Dorsal profile of T13 head capsule showed a weak depression in the middle. Its rostrum was long, cone-shaped and the size was longer than head capsule. T13 had 14 yellowish brown antennal segments and ten abdominal segments. Based the morphological characters, T13 was identified as Havilanditermes atripennis (Haviland).
Based on the morphometric identification, a total of 13 species of termite from six different genera were found in Sarawak Oil Palm Plantation peat zone (Fig. 7). Genus of Schedorhinotermes were the most diverse in term of species (4 spp. or 31%), followed by Nasutitermes (3 spp. or 23%), Coptotermes (2 spp. or 15%), Pericapritermes (2 spp. or 15%), Havilanditermes (1 sp. or 8%) and Parrhinotermes (1 sp. or 8%).
This study found that species of Coptotermes are one of the most commonly encountered species in oil palm plantation which is established on former peat land. Coptotermes curvignathus was abundantly found in the studied site. This is because the species is known to have voracious appetite for living trees instead of dead or decayed wood. The oil palm plantation provides a generous feeding ground for this species. In contrast, Coptotermes was less dominant in rural area where termite infestation was reported (Kirton, 2005). The other species of Coptotermes, C. gestroi which is responsible for most of the urban dwelling in Malaysia (Kirton and Brown, 2003) was not found in the study areas.
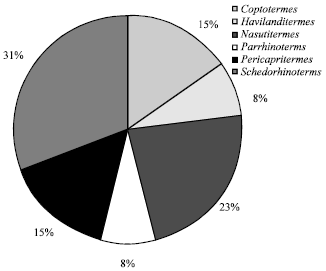 | |
| Fig. 7: | Termite diversity in a Sarawak oil palm peat zone based on genus |
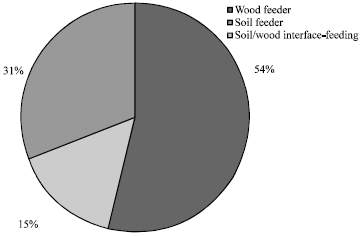 | |
| Fig. 8: | Termite diversity in a Sarawak oil palm peat zone based on feeding group |
All termites that were found in the study sites were exclusively from Rhinotermitidae and Termitidae families. Rhinotermitidae was outnumbering species of Termitidae in the study areas. This is in contrast with Eggleton (2000) that studied on termite assemblage in a conserved Dipterocarp forest in Sabah. They reported that the Rhinotermitidae made up only moderate proportion of the whole assemblages in dipterocarp forest and the most dominant termite fauna were the termes-group soil feeders and Nasutitermes. The contrast findings between Eggleton (2000) and ours showed that agricultural practices may have some effect on the termite assemblages especially on the wood feeder termites as well as Termitidae species which are also known as higher termites. In our study, the wood feeders constitute 54% of the total termites found in the studied areas (Fig. 8), outnumbering the soil and soil/wood feeders. These may be due to changes in soil properties as consequences of agricultural practices which often have numerous repercussions on the physical and biological properties on soil. Studies have shown that the species varieties and abundance of soil microbes in tropical agricultural lands is less than 50% of that of the primary forest originally on the land (Matson et al., 1997) and soil carbon can drop more than 50% within five years when tropical soils were converted for agricultural purposes. In addition to these, when tropical soils are cultivated, they become substantially compacted and surface albedo is increased leading to temperature increase and decreases in precipitation. These all in all may have transformed the ecosystem into a hostile environment for certain species of termites especially species of soil termites and higher termites which often depending on soil microbes or fungi.
The result also showed that lesser negative impact of agricultural practices on wood feeding termites as the cultivation provides food to wood feeder termites. Rhinotermitidae species which often feed on economic importance crops (Edwards and Mill, 1986; French et al., 1986; Su and Scheffrahn, 1998) hence was observed to have the biggest biomass density in this studied agrisystem. However, the faunistics difference before the cultivation intervention between this study site and conserved dipterocarp forest should be taken note of as both sites are in Sunda region.
CONCLUSION
The result of this study has shown that the wood feeder termites are the main species in oil palm plantation that is situated on former peat swamp. The major pest is C. curvignathus which infests living agricultural plants. Scavengers such as Schedorhinotermes spp., Pericapritermes spp., H. atripennis, C. sepangensis and P. aequalis were commonly found in oil palm plantation however, they induce no economical damage to oil palm trees and probably may have positively improved soil condition. These findings suggest that chemical termiticides draining which is a common practice in oil palm plantations may be unnecessary. This is because the practice may negatively affect the termites’ assemblages in agricultural land and causes unnecessary ecology drift. Baiting system using rubber wood chunk to attract C. curvignathus will be a better approach and cause less damage to soil and higher termites.
ACKNOWLEDGMENTS
This research was supported by the RUGS grant (Grant No. 91969 to Patricia King Jie Hung) from Universiti Putra Malaysia, IRPA grant (Grant No. 09-02-02-2008-BTK/ER-36 to Nor Muhammad Mahadi) from the Ministry of Science, Technology and Environment, Malaysia and e-Science grant (Grant No. 05-01-04-SF0516 to Joseph Choon Fah Bong) from the Ministry of Science Technology and Innovations, Malaysia. The authors would like to thank Wong Chi Kang for his their technical help.
| REFERENCES | |
| Donovan, S.E., P. Eggleton and D.E. Bignell, 2001. Gut content analysis and new feeding group classification of termites. Ecol. Entomol., 26: 356-366. Direct Link | Edwards, R. and A.E. Mill, 1986. Termites in buildings: Their biology and control. East Grinstead, Rentokil Ltd., UK. Eggleton, P., 2000. Global Patterns of Termite Diversity. In: Termites: Evolution, Sociality, Symbioses, Ecology, Abe, T., D.E. Bignell and M. Higasi (Eds)., Kluwer Academic Publishers, Dordrecht, pp: 25-51. French, J.R.J., R.J. Robinson and D.M. Ewart, 1986. Mound colonies of Coptoternies lacteus (Isoptera) eat cork in preference to sound wood. Sociobiology, 11: 303-309. Jones, C.G., J.H. Lawton and M. Shachak, 1994. Organisms as ecosystem engineers. Oikos, 69: 373-386. Jones, D.T. and P. Eggleton, 2000. Sampling termite assemblages in tropical forests: Testing a rapid biodiversity assessment protocol. J. Applied Ecol., 37: 191-203. CrossRef | Direct Link | Kambhampati, S. and P. Eggleton, 2000. Phylogenetics and Taxonomy. In: Termites: Evolution, Sociality, Symbioses, Ecology, Abe, T., D.E. Bignell and M. Higasi (Eds.). Kluwer Academic Publishers, Dordrecht, pp: 1-23. Kirton, I.G., V.K. Brown and M. Azmi, 1999. The pest status of the termite, Coptotermes curvignathus, in Acacia mangium plantations: Incidence, mode of attack and inherent predisposing factors. J. Trop. For. Sci., 11: 822-831. Kirton, L.G. and V.K. Brown, 2003. The taxonomic status of pest species of Coptotermes in Southeast Asia: Resolving the paradox in the pest status of the termites, Coptotermes gestroi, C. havilandi and C. travians (Isoptera: Rhinotermitidae). Sociobiology, 42: 43-63. Direct Link | Kirton, L.G., 2005. The importance of accurate termite taxonomy in the braoder perspective of termite management. Proceedings of the 5th International Conference on Urban Pests, July 11-13, 2005, Singapore, pp: 1-7. Lavelle, P., D. Bignell, M. Lepage, V. Wolters and P. Roger et al., 1997. Soil function in a changing world: The role of invertebrate ecosystem engineers. Eur. J. Soil Biol., 33: 159-193. Direct Link | Lawton, J.H., 1996. The Role of Species in Ecosystems: Aspects of Ecological Complexity and Biological Diversity. In: Biodiversity: An Ecological Perspective, Abe, T., S.A. Levin and M. Higashi (Eds.). Springer, New York, USA., pp: 215-228. Matson, P.A., W.J. Parton, A.G. Power and M.J. Swift, 1997. Agricultural intensification and ecosystem properties. Science, 277: 504-509. CrossRef | Direct Link | Prestwich, G.D., 1984. Defense mechanisms of termites. Annu. Rev. Entomol., 29: 201-232. CrossRef | Pringle, R.M., D.F. Doak, A.K. Brody, R. Jocque and T.M. Palmer 2010. Spatial pattern enhances ecosystem functioning in an African Savanna. PLoS Biol., 8: e1000377-e1000377. CrossRef | Stork, N.E. and P. Eggleton, 1992. Invertebrates as determinants and indicators of soil quality. Am. J. Alternative Agr., 7: 38-47. CrossRef | Su, N.Y. and R.H. Scheffrahn, 1998. A review of subterranean termite control practices and prospects for integrated pest management programmes. Integr. Pest Manage. Rev., 3: 1-13. Tartar, A., M.M. Wheeler, X. Zhou, M.R. Coy, D.G. Boucias and M.E. Scharf, 2009. Parallel metatranscriptome analyses of host and symbiont gene expression in the gut of the termite Reticulitermes flavipes. Biotechnol. Biofuels, Vol. 2. 10.1186/1754-6834-2-25 Thapa, R.S., 1981. Termites of Sabah (East Malaysia). Sandakan: Sabah Forest Department, Malaysia. Tho, Y.P. and L.G. Kirton, 1992. The economic significance of Coptotermes termites in Malaysian forestry. Proceedings of the 3rd International Conference on Plant Protection in the Tropics, March 20-23, 1990, Malaysian Plant Protection Society, Kuala Lumpur, Malaysia, pp: 193-199. Tho, Y.P., 1992. Termites of Peninsular Malaysia. Forest Research Institute Malaysia, Malaysia, Kepong, Malaysia. Turner, J.S., 2004. Gaia, Extended Organisms and Emergent Homeostasis. In: Scientists Debate Gaia: The Next Century, Schneider, S.H., J.R. Miller, E. Crist and P.J. Boston (Eds). The MIT Press, Cambridge, pp: 57-70. Warnecke, F., P. Luginbuhl, N. Ivanova, M. Ghassemian and T.H. Richardson et al., 2007. Metagenomic and functional analysis of hindgut microbiota of a wood-feeding higher termite. Nature, 450: 560-565. CrossRef | PubMed | Direct Link | |
For further details log on website :
http://scialert.net/fulltext/?doi=je.2012.68.78&org=10









No comments:
Post a Comment