Published Date
Author
 (1)
(1)



For further details log on website :
http://journals.plos.org/plosone/article?id=10.1371/journal.pone.0125376
- Published: May 15, 2015
- http://dx.doi.org/10.1371/journal.pone.0125376
Author
Abstract
Coptotermes formosanus is an imported, subterranean termite species with the largest economic impact in the United States. The frontal glands of the soldier caste termites comprising one third of the body mass, contain a secretion expelled through a foramen in defense. The small molecule composition of the frontal gland secretion is well-characterized, but the proteins remain to be identified. Herein is reported the structure and function of one of several proteins found in the termite defense gland secretion. TFP4 is a 6.9 kDa, non-classical group 1 Kazal-type serine protease inhibitor with activity towards chymotrypsin and elastase, but not trypsin. The 3-dimensional solution structure of TFP4 was solved with nuclear magnetic resonance spectroscopy, and represents the first structure from the taxonomic family, Rhinotermitidae. Based on the structure of TFP4, the protease inhibitor active loop (Cys8 to Cys16) was identified.
Figures
Citation: Negulescu H, Guo Y, Garner TP, Goodwin OY, Henderson G, Laine RA, et al. (2015) A Kazal-Type Serine Protease Inhibitor from the Defense Gland Secretion of the Subterranean Termite Coptotermes formosanus Shiraki. PLoS ONE 10(5): e0125376. doi:10.1371/journal.pone.0125376
Academic Editor: Ben J. Mans, Onderstepoort Veterinary Institute, SOUTH AFRICA
Received: October 15, 2014; Accepted: March 24, 2015; Published: May 15, 2015
Copyright: © 2015 Negulescu et al. This is an open access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited
Data Availability: All relevant structural data are available from the Protein Data Bank (id 2N17) and the Biological Magnetic Resonance Data Bank (accession number 18896).
Funding: RAL and GH received funding from the United States Department of Agriculture-Agricultural Research Service New Orleans, specific cooperative agreement 58-6435-8-084, (http://www.ars.usda.gov/main/site_main.htm?modecode=64-35-00-00).
Competing interests: The authors have declared that no competing interests exist.
Introduction
Termite activity has an annual economic impact of over 1 billion dollars in the United States [1,2]. This impact is caused by termite consumption of wood fiber (from live trees and wood structures), crops, plants, and other materials, which contain cellulose. The most destructive and economically important genera are Coptotermes and Reticulitermes of the subterranean termite family, Rhinotermitidae. Members of the imported Coptotermitidae subfamily are very active in the southern states [1,3].
A termite colony contains a soldier caste, which uses its frontal defense gland secretion and pincer-modified mandibles as weapons to protect the colony. In a Coptotermes formosanuscolony, soldiers comprise 5–28% of the entire colony and up to 60% of its foraging members [4–6]. The defense gland is well developed in Rhinotermitidae and Termitidae (desert termite) and is a cephalic organ that opens through a frontal pore or fontanelle [7]. The gland reaches deep into the abdominal cavity, and the secretion can comprise as much as one third of the total body weight [8]. During combat, the milky defense secretion of its frontal gland is ejected through a frontal pore onto the attacker, rapidly stiffening in the air [7]. Although much work has been done to determine the chemical composition of the frontal gland secretion from different families of termites, the protein components of the secretions is relatively overlooked.
Henderson, Laine, and others have identified various components of the frontal gland secretion in Coptotermitinae, including n-alkanes, mucopolysaccharide [9], naphthalene [10], free fatty acids, hexacosanoic acid and lignoceric acid [11], and novel ceramides [12]. Recent work has generated interest in caste-specific gene expression. SOL1 is a protein that is found only in the mandibular glands of Hodotermopsis japonica, but not in any other stages of differentiation into presoldiers [13]. Ntsp1 is a secretory carrier protein present only in the epithelial cells of the frontal gland reservoir of Nasutitermes takasagoensis soldiers [14]. In Reticulitermes flavipes, the most common termite in the United States, Scharf et al. describe the presence of soldier-specific gene expression of transcription and translation factors with significant sequence homology to the bicaudal and bric-a-brac genes in Drosophila, which function in embryonic pattern formation [15]. During the research, seven proteins were identified using polyacrylamide gel electrophoresis and amino acid sequencing in the defensive secretion of the frontal gland of C. formosanus soldiers. Two proteins had lipocalin homologies. A third, novel protein, named TFP4, has homology to Kazal-type serine protease inhibitors and, due to its potential for determining activity and its small size, is amenable to NMR analysis; it is the first of the secretion proteins to be characterized.
Kazal-type serine protease inhibitors are small proteins, 40–60 amino acids in length, with a structural fold constrained by three disulfide bonds arranged in the order: I-V, II-IV, and III-VI [16,17]. Kazal-type protease inhibitors are classified as “classical” or “non-classical” (group 1 or group 2) based on the relative positions of cystines I and V in the sequence [18,19]. The structures also contain a central α-helix, three-strand anti-parallel β-sheet, and reactive site loop between cystines II and III [16]. The reactive site loop is exposed from the protein's globular structure and adopts a conformation that is complementary to the protease surface [20]. For most classical and non-classical Kazal-type protease inhibitors, the reactive loop, including cystines II and III, is comprised of 9 amino acid residues with little consensus (S1 Fig) [17]. Using nomenclature proposed by Laskowski, et al. [21], the third amino acid in the loop sequence, P1, is the reactive site residue of the protease inhibitor and determines the enzyme’s specificity (Fig 1). Kazal-type serine protease inhibitors with lysine and arginine at the P1 site inhibit trypsin-like enzymes; those with tyrosine, phenylalanine, leucine and methionine inhibit chymotrypsin-like enzymes; and those with alanine, serine, leucine, and methionine at the reactive P1 site inhibit elastase-like enzymes [22,23]. The reactive site is typically encompassed by at least one disulfide bridge [24]. Prolines at residues P2 and P4’ help to ensure proper reactive site geometry, but are not necessary [21]. The Kazal-type protease inhibitors have a similar structural fold, but little sequence consensus, affording the possibility of dual-functionality. Here are presented the activity assays and 3-dimensional structure of TFP4. To our knowledge, TFP4 is the first protein characterized from the termite defense gland secretion
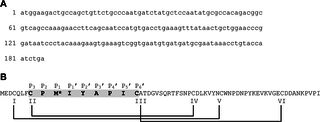
Fig 1. The (A) DNA and (B) primary amino acid sequence of TFP4 cloned from the defense gland secretion of C. formosanus termite soldiers.
The Kazal-type serine protease inhibitor consensus sequence is highlighted in grey with the active site residue, P1, indicated with the asterisk at Met10. Laskowski and Kato’s nomenclature for the Kazal-type active loop is shown above the amino acid sequence [21]. Cystines and disulfide bonds are indicated by Roman numerals and lines.
Materials and Methods
Crude protein extract collection
Between 150–200 healthy C. formosanus soldiers were collected from colonies in Brechtel Park, New Orleans, Louisiana. No specific permissions or field permits were required for this location and activity, and the field studies did not involve endangered or protected species. The reason no specific permissions were required is because termites are not a desired insect to have in parks where they are damaging trees. Coptotermes formosanus is an invasive species that was brought over during WWII as ships in the Pacific transported infested materials from Japanese islands to port cities in South Carolina, Texas, and Louisiana. They are native to China where they remain the most important pest of wood structures and live trees. Collections of termites have been made from Brechtel Park in New Orleans since 1999. A crate full of wood is buried in the ground near a heavily infested tree and brought to the lab after a month containing up to 30,000 individuals at a time. The termites are contained with food until needed for experiments. The frontal defense gland secretion was isolated on ice by squeezing the head near the frontal pore with tweezers. The secretion was transferred into 5 ml of buffer (10 mM phosphate, 0.15 M sodium chloride, pH 7.0) and centrifuged at 5000 x g for 10 min. The supernatant fraction was filtered (2 μm), dialyzed for 48 hr (3000 Da molecular weight cut-off), and lyophilized.
Isolation of total mRNA from C. formosanus soldiers and cDNA library construction
One gram of soldier tissue was prepared for isolation of total mRNA using the Fast Track 2.0 kit (BD Biosciences Clontech) according to the protocol provided by the manufacturer. For the construction of the cDNA library, the BD SMART (Switching Mechanism At 5’ end of RNA Transcript) RACE (Rapid Amplification of cDNA Ends) kit for cDNA synthesis from BD Biosciences Clontech was used by following the manufacturer’s protocol. Oligonucleotides used for the cDNA library construction include: RT reaction primer: 5’-AAGCAGTGGTATCAACGCAGATAC(T)30VN-3’ (V = A, G, or C), and cDNA library construction primers: 5’-GAYTGBCARCTNTTYTGBCCNATG-3’ (where Y = T or C, B = T, C, or G; R = A or G), 5’-CTAATACGACTCACTATAGGGCAAGCAGTGGTATCAACGCAGAGT-3’, 5’-CTAATACGACTCACTATAGGGC-3’ (where V = A, G or C). PCR reactions were performed using a thermal cycler, and PCR purification was performed using the QIAquick PCR Purification kit using the reagents and protocol provided.
The purified PCR product was sequenced by the GeneLab sequencing facility at Louisiana State University. A search of the public nucleotide and protein database was performed using the BLAST software [25,26] provided by the National Center for Biotechnology Information. MUSTER (Multi-Sources ThreadER) [27] was used to identify template structures from the Protein Data Bank (PDB) [28] using the TFP4 amino acid sequence. The output from MUSTER includes full-length models of TFP4 built by MODELLER [29] using the template alignments. The threaded structures were visualized with UCSF Chimera software [30].
Cloning and protein expression
The cDNA of interest was cloned using the pET46Ek/LIC system (Novagen), which provided an N-terminal 6X-His tag followed by an enterokinase site (MAHHHHHHVDDDDKP). Oligonucleotides used: 5’-GACGACGACAAGATAGAAGACTGCCAGCTGTT-3’ and 5’-GCTACACCAGCAACATCAACCGGGATTCTCCTC-3’. The recombinant plasmid was transformed into E. coli BL21(DE3) cells from Novagen for protein expression. The cells were grown in LB media to produce protein for the activity assays and minimal media to produce isotopically labeled protein for NMR analysis. The 15N/13C-M9 minimal media contained 13 g/L Na2HPO4, 3 g/L KH2PO4, 0.5 g/L NaCl, 1 g/L 15NH4Cl (Cambridge Isotope Laboratories), 2 g/L uniform 13C-glucose (Cambridge Isotope Laboratories), 2 mM MgSO4, 0.1 mM CaCl2, vitamins, and trace metals. The cells were grown at 37°C to an optical density at 600 nm of 0.8, induced with 1 mM isopropyl β-D-1-thiogalactopyranoside, and grown at 37°C for 4 hours. TFP4 was purified from the cleared lysate using nickel or cobalt affinity media and gel filtration chromatography.
Protease inhibition assays
Elastase (EC 3.4.21.26) and N-succinyl-(L-alanine)3-p-nitroanilide were purchased from the Sigma-Aldrich Co. Complete Protease Inhibitor cocktail was purchased from Roche. The assays and enzyme activity calculations were performed according to the manufacturer’s protocol, based on the method of Bieth et al. [31]. The reactions were monitored at 410 nm at room temperature using a spectrophotometer.
Chymotrypsin (EC 3.4.21.1) and N-benzoyl-L-tyrosine ethyl ester (BTEE) were purchased from Worthington Biochemical Corp. and Acros Organics, respectively. The assays to measure enzyme activity and reaction velocity were performed according to the method of Humel et al. [32]. The change in light absorbance at 256 nm was monitored at room temperature using a spectrophotometer. The reaction velocity was converted to units of mM/min using the molar extinction coefficient of N-benzoyl-L-tyrosine (0.964 mM-1min-1). The inhibition constant (Ki) of TFP4 was determined using samples at 50, 75, 100, 200, and 300 μM BTEE with 0, 250, and 500 nM of recombinantly expressed and purified TFP4. Initial velocities (V) at each substrate (S) and inhibitor (I) concentration were analyzed by nonlinear regression analysis using Origin software. The data were globally fitted to competitive, noncompetitive, and uncompetitive inhibition models of which the best model was competitive inhibition (Eq 1). The Ki, standard error, and correlation coefficient (R2) were determined from the best global fit to Eq 1, where Vmax is the maximum reaction velocity and Km is the Michaelis-Menten constant.
Trypsin (EC 3.4.21.4) and Nα-benzoyl-L-arginine ethyl ester were purchased from the Sigma-Aldrich Co. The assays and enzyme activity calculations were performed according to the method described by Bergmeyer et al. [33]. Changes in light absorbance were measured at 253 nm at room temperature using a spectrophotometer.
Protein NMR analysis and structure refinement
All NMR data was collected on a Varian VS-700 MHz spectrometer equipped with a 5-mm HCN-5922 cold probe in the Louisiana State University, Department of Chemistry, NMR facility. 15N,13C-TFP4 was concentrated to 1 mM in buffer (50 mM potassium phosphate, 50 mM NaCl, 10% deuterated water, pH 6.5) for NMR structure determination. Resonance assignments (1H, 13C, and 15N) were determined using conventional triple-resonance NMR methods [34], the program CcpNmr Analysis [35], and by manual assignment. Chemical shifts have been deposited in the Biological Magnetic Resonance Data Bank (BMRB accession number 18896) [36]. Structure calculations were performed using the program, Crystallography & NMR System (CNS) version 1.2 [37], based on 1H-1H NOE derived distance constraints calculated from peak volumes using the CcpNmr Analysis program [35], backbone dihedral angle constraints derived from chemical shifts using the programs, CcpNmr Analysis and TALOS+ [38], and disulfide bonds at Cys4-Cys37, Cys8-Cys30 and Cys16-Cys52. The existence of disulfide bonds was confirmed by the 13C Cβ chemical shifts of the oxidized cysteine residues [39]. The configuration of the disulfide bonds, based on other Kazal-type protease inhibitor structures, was verified by NOE constraints between Cys8-Cys30 and Cys16-Cys52. Validation of the resulting 10 refined conformers was performed with the Protein Structure Validation Software (PSVS) server version 1.5 [40], including structural statistics and global structure quality factors: Verify3D [41], ProsaII [42], PROCHECK [43], and MolProbity [44]. The final refined ensemble of 10 models and NMR constraint data have been deposited in the PDB (ID 2N17) [28]. Molecular graphics images and protein alignments were produced using the UCSF Chimera software [30]. The average structure, based on the 10 models, was calculated using CNS [10] and analyzed with the Dali server [45] to identify structural homologs in the PDB.
Results
Molecular cloning of TFP4 protein from C. formosanus soldiers defense gland secretion
Sodium dodecylsulfate polyacrylamide gel electrophoresis (SDS-PAGE) of the C. formosanussoldiers’ defense gland secretion revealed the existence of seven proteins. N-terminal sequence determination was performed and one protein, named TFP4, was chosen for further characterization. A cDNA library was constructed using total mRNA from the tissue. Based on the available N-terminal sequence, the corresponding cDNA was isolated. The cloned cDNA was sequenced and included a 186 bp reading frame encoding 61 amino acids with a calculated molecular mass of 6888 Da (Fig 1). The nucleotide sequence had no significant alignments using BLAST [25,26], and the protein sequence showed moderate homology to a salivary gland protease inhibitor from Nauphoeta cinerea, a cockroach species found in tropical climates. This homology is not surprising since termites and cockroaches descend from a common ancestor. The results showed 40% sequence identity, 54% sequence similarity, and a length of 72 amino acids, similar to TFP4’s 61 amino acids. The cDNA of TFP4 was cloned into a pET46Ek/LIC vector (Novagen) for expression in E. coli. An N-terminal 6X-His tag was included in the recombinant sequence for protein purification (S1 Fig).
TFP4 is a Kazal-type serine protease inhibitor
Amino acid sequence analysis and threading identified a Kazal-type domain at the N-terminus of TFP4 (residues 3–37, Fig 1B). The top five template structures that aligned with the TFP4 sequence using MUSTER (Multi-Sources ThreadER) [27] were all Kazal-type inhibitors: Anemonia elastase inhibitor (PDB ID 1Y1B, chain A) [28,46], Triatoma infestans factor XIIa inhibitor (infestins 4, PDB ID 2ERW) [47], Rhodnius prolixus thrombin inhibitor (rhodniin, PDB ID 1TBQ, chains S and R) [48], Dipetalogaster maximus thrombin inhibitor (dipetalin, PDB ID 1KMA) [49], and the third domain of turkey ovomucoid inhibitor in complex with alpha-chymotrypsin (PDB ID 1HJA) [50]. Three of the five structures belong to insect proteins—T. infestans, R. prolixus, and D. maximus—all hematophagous insects with inhibitors to prevent blood coagulation [47–49]. The other two structures from Anemonia and turkey are serine protease inhibitors for elastase and alpha-chymotrypsin, respectively [46,50]. The structures predict that TFP4 has a Kazal-type domain with an active loop at amino acids 8–16, Met10 at the active site position P1 (Fig 1B), an α-helix, a three strand anti-parallel β-sheet, and three disulfide bonds (Cys4-Cys37, Cys8-Cys30 and Cys16-Cys52).
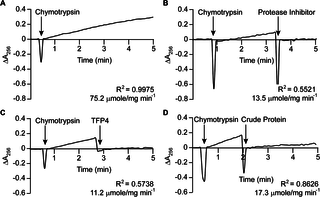
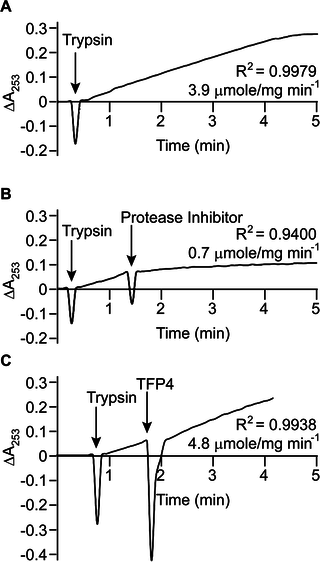
Kazal-type enzymes inhibit serine proteases, a group of proteins which includes trypsin, chymotrypsin, and elastase. TFP4 inhibition was tested by monitoring hydrolysis rates of chromophoric substrates in the presence of each protease. Negative controls of the protease only and positive controls with protease inhibitor cocktail from Roche (60 μg/mL) were performed. Protease inhibition by TFP4 (2 μg/mL) and crude protein extract from the C. formosanus frontal gland secretion (1.2 μg/mL) were tested with elastase (Fig 2), chymotrypsin (Fig 3), and trypsin (Fig 4). The spectrophotometric traces in Figs 2, 3, and 4 show the hydrolysis rate and correlation coefficient calculated for each reaction. The baseline hydrolysis rates of elastase, chymotrypsin, and trypsin with their respective substrates were 10.2 ± 0.3 (2 replicates), 75.2 ± 1.6 (3 replicates), and 3.9 ± 0.2 (3 replicates) μmoles/mg min-1, respectively. As expected, the Roche protease inhibitor cocktail decreased the hydrolysis rate for each protease. Recombinant TFP4 and the crude protein extract showed inhibition of elastase and chymotrypsin, but not trypsin. TFP4 decreased the hydrolysis rate of elastase to 0.7 μmole/mg min-1 and chymotrypsin to 11.2 μmole/mg min-1. For trypsin, the hydrolysis rate with TFP4 (4.8 μmoles/mg min-1) did not change from the baseline rate.
Fig 2. Elastase inhibition assays.
(A) Baseline elastase (0.2 μg/mL) activity with N-succinyl-(l-alanine)3-p-nitroanilide (0.27 mM). Inhibition of elastase with (B) Roche protease inhibitor cocktail (60 μg/mL), (C) TFP4 (2 μg/mL), and (D) crude protein extract (1.2 μg/mL) from the defense gland secretion of C. formosanus. Additions are indicated by arrows. The hydrolysis rate and correlation coefficient (R2) are shown for each reaction.
Fig 3. Chymotrypsin inhibition assays.
(A) Baseline chymotrypsin (0.1 μg/mL) activity with N-benzoyl-L-tyrosine ethyl ester (0.5 mM). Inhibition of chymotrypsin with (B) Roche protease inhibitor cocktail (60 μg/mL), (C) TFP4 (2 μg/mL), and (D) crude protein extract (1.2 μg/mL) from the defense gland secretion of C. formosanus. Additions are indicated by arrows. The hydrolysis rate and correlation coefficient (R2) are shown for each reaction.
Fig 4. Trypsin inhibition assays.
(A) Baseline trypsin (0.3 μg/mL) activity with Nα-benzoyl-L-arginine ethyl ester (75 μM). Inhibition of trypsin with (B) Roche protease inhibitor cocktail (60 μg/mL) and (C) TFP4 (2 μg/mL). Additions are indicated by arrows. The hydrolysis rate and correlation coefficient (R2) are shown for each reaction. The assay with crude extract was not performed since no inhibition was observed with the recombinant protein.
The Ki of TFP4 for chymotrypsin was determined by measuring the hydrolysis rate of the chromophoric substrate, BTEE, at varying BTEE and TFP4 concentrations. The data shown in Fig 5 were globally fit to a competitive inhibition model to give a Ki of 91.3 ± 8.9 nM TFP4. This very strong, competitive inhibition by TFP4 for chymotrypsin is similar to other Kazal-type protease inhibitors [17].
Fig 5. Enzyme kinetic analysis of TFP4 inhibition of BTEE hydrolysis by chymotrypsin.
The reaction velocities of chymotrypsin hydrolysis of BTEE at varying concentrations of BTEE and TFP4 are plotted. The points are the observed velocities, and the lines represent the best global fit of the data to Eq 1, a competitive inhibition model. The Ki of TFP4 determined from the fit (91.3 ± 8.9 nM) and the correlation coefficient (R2 = 0.9884) are displayed on the plot.
Solution structure of TFP4 by NMR
The solution NMR structure of TFP4 was determined using triple-resonance NMR and NOESY experiments. An assigned 1H,15N heteronuclear single quantum coherence spectrum is shown in S3 Fig, and structural statistics are summarized in S1 Table. The structure was determined using 626 constraints including 526 NOE-based distance constraints (100 long range NOE constraints), and 100 dihedral angle constraints [38]. Structure refinement was performed using CNS [37], and the 10 lowest energy structures from 100 structures were selected. The average backbone root mean square deviation (RMSD) of ordered residues in the final ensemble is 0.4 Å (S4 Fig). Backbone dihedral angle analysis indicated that 100% fall in the most favored or allowed regions of the Ramachandran plot [44]. Overall, the global structure quality factors indicate a good, solution structure of TFP4. The average structure in Fig 6 shows an α-helix (Pro29-Trp38) and a three-strand anti-parallel β-sheet (Ile15-Ala17, Arg24-Phe26, Glu46-Lys48). NOE constraints support the predicted disulfide bond arrangement of Cys4-Cys37, Cys8-Cys30, and Cys16-Cys52. Chemical shifts have been deposited in the Biological Magnetic Resonance Data Bank (BMRB accession number 18896) [36]. The final refined ensemble of 10 models and constraint lists have been deposited in the PDB (ID 2N17) [28].
Fig 6. NMR solution structure of TFP4.
Two ribbon views of TFP4 are shown to illustrate the Kazal-type consensus loop. The amino acids are numbered based on the TFP4 sequence in Fig 1B. The asterisks indicate the active site amino acid, Met10. The secondary structure is colored from the N- to C-terminus, blue to red. The disulfide bonds are labeled with Roman numerals to indicate the cystine connectivity. Note that partial transparency of the second β-sheet is used in the structure on the right to show the aft III-VI disulfide bond.
Discussion
The amino acid sequence, activity, and structure of TFP4 confirm that it is a non-classical group 1 Kazal-type serine protease inhibitor with activity toward elastase and chymotrypsin, but not trypsin. TFP4 has 61 amino acids and a molecular weight of 6888 Da. The consensus sequence for the active site loop (Cys8 to Cys16) has a methione (Met10) in the reactive P1 site, which predicts TFP4’s specificity for chymotrypsin, elastase, and not trypsin [21], corroborating results of the functional assays. The 3D solution structure of TFP4 confirms the non-classical group 1 Kazal-type protease inhibitor structure with three disulfide bonds, a central α-helix, a three-strand anti-parallel β-sheet, and the reactive site loop exposed at the surface (Fig 6) [16]. The position of the disulfide bonds, Cys4-Cys37, Cys8-Cys30, and Cys16-Cys52, classifies TFP4 as a non-classical group 1 Kazal-type with cystines I and V near the C-terminus of the α-helix (Fig 6) [46]. Other structural conditions for Kazal-type protease inhibitor activity are met with two disulfide bridges at either end of the reactive loop (Fig 6) [24] and prolines at residues P2and P4’ to ensure proper reactive site geometry (Fig 1B) [21].
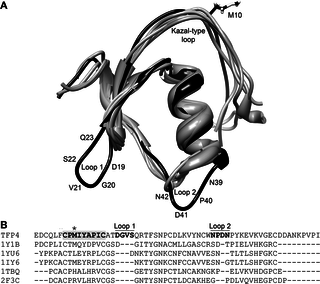
The top five, non-redundant, structural homologs of TFP4, as determined by the Dali server [45], are all Kazal-type inhibitors: anemonia elastase inhibitor [46], turkey ovomucoid third domain [51], silver pheasant ovomucoid third domain [52], Rhodnius prolixus rhodniin [48], and Triatoma infestans infestins [47] (PDB ID: 1Y1B chain A, 1YU6 chain D, 1IY6 chain A, 1TBQ chain S, 2F3C chain I, respectively). A structural and sequence alignment of TFP4 with the homologous structures show loops that are not present in the other Kazal-type protease inhibitors (Fig 7). The sequence between cystines V and VI is not well conserved among Kazal-type protease inhibitors and can vary in lengths from 9–21 amino acids [17]. Thus, the extra loop with Asp41 (Loop 2 in Fig 7) is not surprising given the loop can be various lengths. The sequence between cystines III and IV is conserved as determined from 85 invertebrate Kazal-type protease inhibitor sequences [17] using WebLogo v3 [53,54] (S5 Fig). The sequence alignment of TFP4 with the consensus sequence indicates that Ser22-Gln23-Arg24 are extra residues that result in an extended β-hairpin (Loop 1 in Fig 7).
Fig 7. TFP4 structure aligned with structural homologs.
(A) The top five, non-redundant, structural homologs as determined by the Dali server [45] aligned with the average TFP4 structure. TFP4 is shown in black and the structural homologs in grey (PDB ID: 1Y1B chain A, 1YU6 chain B, 1IY6 chain A, 1TBQ chain S, 2F3C chain I) [46–48,51,52]. The extra loops in TFP4 that are not found in the homologous structures are labeled, Loop 1 and Loop 2, with their corresponding amino acid sequences. (B) Sequence alignments based on the structural alignment of TFP4 and the homologous proteins were generated using the Dali server [45]. The Kazal-type consensus sequence is highlighted in bold with the active site residue indicated by the asterisk. Loops 1 and 2 are labeled with the corresponding residues are in bold.
TFP4 is the first protein cloned and characterized from the frontal defense gland secretion of termite soldiers. TFP4’s function is likely to protect the content of the termite defense gland in C. formosanus from microbial proteolytic enzymes. This gland has contents that comprise one third of the soldier’s mass with a foramen in the head that could conceivably be accessed by bacteria and fungi. The protease inhibitor activity may be antimicrobial as bacteria and fungi produce proteases during the infection process [55,56]. The protease activity may also assist in the protection of the protein content of the defense fluid, whose additional functions remain to be determined. The 3D solution structure of TFP4 is the first structure deposited in the PDB (ID 2N17) from the family Rhinotermitidae (subterranean termites) and only the fourth protein from the order Isoptera (termites) [57–60].
Supporting Information
S1 Fig. The (A) sequence logo for the Kazal-type protease inhibitor active loop and (B) its alignment with the TFP4 sequence.
(A) The logo was created using 83 invertebrate Kazal-type protease inhibitors [17] for the amino acid sequence between cystines II and III using WebLogo v.3 [53]. The overall height at each position indicates the relative sequence conservation, and the heights of the symbols indicate the relative frequency of each amino acid. (B) The asterisks indicate agreement between the consensus sequence and TFP4. Laskowski and Kato’s nomenclature for the Kazal-type protease inhibitor sequence is shown [21].
doi:10.1371/journal.pone.0125376.s001
(TIF)
S2 Fig. The recombinant TFP4 sequence with a 6X-His tag.
The 6X-His tag is underlined with numbering from (-15 to -1). The TFP4 sequence starts at Glu2 and is numbered to be consistent with Fig 1B.
doi:10.1371/journal.pone.0125376.s002
(TIF)
S3 Fig. An assigned 1H-15N heteronuclear single quantum coherence spectrum of 15N,13C-TFP4.
Assignments of the amide backbone peaks are indicated by the residue number and single character amino acid code next to each peak. Dashed lines indicate side-chain peaks. Residues are numbered as in the S2 Fig.
doi:10.1371/journal.pone.0125376.s003
(TIF)
S4 Fig. Ensemble of 10 NMR structures of TFP4.
The 10 lowest energy structures from 100 models were aligned using the PSVS server [40]. Residues Glu2-Ile61 without the 6X-His tag are shown in ribbon view with the disulfide bonds in black. The RMSD of the ordered residues in the 10 models (Cys4-Val59, defined by conformationally restricting NMR constraints) is 0.4 Å for the backbone heavy atoms and 0.9 Å for all heavy atoms.
doi:10.1371/journal.pone.0125376.s004
(TIF)
S5 Fig. The sequence conservation of (A) the 12 amino acids between cystines III and IV of invertebrate Kazal-type domains compared to (B) the corresponding TFP4 sequence.
WebLogo v.3 [53] was used to create the consensus sequences using 85 invertebrate sequences. The overall height at each position indicates the relative sequence conservation, and the heights of the symbols indicate the relative frequency of each amino acid. (B) The asterisks indicate agreement between the TFP4 sequence and the consensus sequences. The periods indicate gaps added to align the two sequences. The dash indicates very low sequence conservation.
doi:10.1371/journal.pone.0125376.s005
(TIF)
S1 Table. NMR and structural statistics.
Structural statistics were computed using the Protein Structure Validation Software (PSVS) Version 1.5 [40] for the ensemble of 10 NMR structures (S4 Fig) deposited in the PDB (ID 2N17) [28]. All analyses were performed using the ordered residues, Cys4-Val59.
doi:10.1371/journal.pone.0125376.s006
(DOCX)
Acknowledgments
We thank Dr. Grover Waldrop and Dr. Richard Galardy for helpful discussions regarding protease inhibitors and enzymology, and Drs. C. Husseneder, J. Beuzelin and W. Cai with Louisiana State University Agricultural Center for their review of this manuscript. Horia Negulescu was the recipient of a Louisiana Board of Regents Economic Development Assistantship and an LSU Graduate School Enhancement Award. This paper is part of his dissertation research, supervised by Roger A. Laine.
Author Contributions
Conceived and designed the experiments: HN YG TPG OYG GH RAL MAM. Performed the experiments: HN YG TPG OYG GH RAL MAM. Analyzed the data: HN YG TPG OYG GH RAL MAM. Contributed reagents/materials/analysis tools: GH RAL MAM. Wrote the paper: HN RAL MAM.
References
- 1.Lax AR, La Osbrink W (2003) United States Department of Agriculture—Agriculture Research Service—research on targeted management of the Formosan subterranean termite Coptotermes formosanus Shiraki (Isoptera: Rhinotermitidae). Pest Manag Sci 59: 788–800. pmid:12846330 doi: 10.1002/ps.721
- 2.Pimentel D, Zuniga R, Morrison D (2005) Update on the environmental and economic costs associated with alien-invasive species in the United States. Ecol Econ 52: 273–288. doi: 10.1016/j.ecolecon.2004.10.002
- 3.Mullins AJ, Su N-Y, Owens C (2011) Reinvasion and colony expansion of Coptotermes formosanus (Isoptera: Rhinotermitidae) after areawide elimination. J Econ Entomol 104: 1687–1697. pmid:22066200 doi: 10.1603/ec11036
- 4.King EG, Spink WT (1975) Development of incipient Formosan subterranean termite colonies in the field. Ann Entomol Soc Am 68: 355–358. doi: 10.1093/aesa/68.2.355
- 5.Nakajima S, Shimizu K, Nakajima Y (1964) Analytical studies of the vitality of colonies of the Formosan termite, C. formosanus Shiraki: II. Seasonal fluctuations on the external characters of the workers, the ratio of caste-number and carbon dioxide in the next of a colony. Miyazaki Univ, Faculty Agric Bull 9: 222–227.
- 6.Su NY, La Fage JP (1999) Forager proportions and caste compositions of colonies of the Formosan subterranean termite (Isoptera: Rhinotermitidae) restricted to cypress trees in the Calcasieu River, Lake Charles, Louisiana. Sociobiology 33: 185–193.
- 7.Prestwich GD (1984) Defense mechanisms of termites. Annu Rev Entomol 29: 201–232. doi: 10.1146/annurev.en.29.010184.001221
- 8.Quennedey A, Deligne J (1975) L'arme frontale des soldats de termites. I. Rhinotermitidae. Insectes Soc 22: 243–267. doi: 10.1007/bf02223076
- 9.Moore BP (1969) Biochemical studies in termites; Krishna K, Weesner FM, editors. York New: Academic Press.
- 10.Chen J, Henderson G, Grimm CC, Lloyd SW, Laine RA (1998) Naphthalene in Formosan subterranean termite carton nests. J Agric Food Chem 46: 2337–2339. doi: 10.1021/jf9709717
- 11.Chen J, Henderson G, Laine RA (1999) Lignoceric acid and hexacosanoic acid: Major components of soldier frontal gland secretions of the Formosan subterranean termite (Coptotermes formosanus). J Chem Ecol 25: 817–824.
- 12.Ohta M, Matsuura F, Henderson G, Laine RA (2007) Novel free ceramides as components of the soldier defense gland of the Formosan subterranean termite (Coptotermes formosanus). J Lipid Res 48: 656–664. pmid:17164223 doi: 10.1194/jlr.m600387-jlr200
- 13.Miura T, Kamikouchi A, Sawata M, Takeuchi H, Natori S, Kubo T, et al. (1999) Soldier caste-specific gene expression in the mandibular glands of Hodotermopsis japonica(Isoptera: Termopsidae). Proc Natl Acad Sci USA 96: 13874–13879. pmid:10570166doi: 10.1073/pnas.96.24.13874
- 14.Hojo M, Morioka M, Matsumoto T, Miura T (2005) Identification of soldier caste-specific protein in the frontal gland of nasute termite Nasutitermes takasagoensis (Isoptera: Termitidae). Insect Biochem Mol Biol 35: 347–354. pmid:15763470 doi: 10.1016/j.ibmb.2005.01.007
- 15.Scharf ME, Wu-Scharf D, Pittendrigh BR, Bennett GW (2003) Caste- and development-associated gene expression in a lower termite. Genome Biol 4: R62. pmid:14519197doi: 10.1186/gb-2003-4-10-r62
- 16.Kazal LA, Spicer DS, Brahinsky RA (1948) Isolation of a crystalline trypsin inhibitor-anticoagulant protein from pancrease. J Am Chem Soc 70: 3034–3040. pmid:18882536doi: 10.1021/ja01189a060
- 17.Rimphanitchayakit V, Tassanakajon A (2010) Structure and function of invertebrate Kazal-type serine proteinase inhibitors. Dev Comp Immunol 34: 377–386. doi: 10.1016/j.dci.2009.12.004. pmid:19995574
- 18.Krejci K, Fritz H (1976) Structural homology of a trypsin-plasmin inhibitor from leeches (Bdellin B-3) with secretory trypsin-inhibitors from mammals. FEBS Lett 64: 152–155. pmid:131707 doi: 10.1016/0014-5793(76)80271-2
- 19.Augustin R, Siebert S, Bosch TCG (2009) Identification of a Kazal-type serine protease inhibitor with potent anti-staphylococcal activity as part of Hydra's innate immune system. Dev Comp Immunol 33: 830–837. doi: 10.1016/j.dci.2009.01.009. pmid:19428484
- 20.Bode W, Huber R (1992) Natural protein proteinase-inhibitors and their interaction with proteinases. Eur J Biochem 204: 433–451. pmid:1541261 doi: 10.1111/j.1432-1033.1992.tb16654.x
- 21.Laskowski M, Kato I (1980) Protein inhibitors of proteinases. Annu Rev Biochem 49: 593–626. pmid:6996568 doi: 10.1146/annurev.bi.49.070180.003113
- 22.Kato I, Kohr WJ, Laskowski M (1978) Regulatory proteolytic enzymes and their inhibitors; Federation of European Biochemical Societies., Magnusson S, editors. Oxford: Pergamon Press.
- 23.Laskowski M Jr, Sealock RW (1971) 11 protein proteinase inhibitors—molecular aspects. In: Paul DB, editor. The Enzymes: Academic Press. pp. 375–473.
- 24.Ozawa K, Laskowsk M (1966) Reactive site of trypsin inhibitors. J Biol Chem 241: 3955–3961. pmid:5950769
- 25.Altschul SF, Gish W, Miller W, Myers EW, Lipman DJ (1990) Basic local alignment search tool. J Mol Biol 215: 403–410. pmid:2231712 doi: 10.1016/s0022-2836(05)80360-2
- 26.Altschul SF, Madden TL, Schäffer AA, Zhang J, Zhang Z, Miller W, et al. (1997) Gapped BLAST and PSI-BLAST: A new generation of protein database search programs. Nucleic Acids Res 25: 3389–3402. pmid:9254694 doi: 10.1093/nar/25.17.3389
- 27.Wu ST, Zhang Y (2008) MUSTER: Improving protein sequence profile-profile alignments by using multiple sources of structure information. Proteins 72: 547–556. doi: 10.1002/prot.21945. pmid:18247410
- 28.Berman HM, Westbrook J, Feng Z, Gilliland G, Bhat TN, Weissig H, et al. (2000) The Protein Data Bank. Nucleic Acids Res 28: 235–242. pmid:10592235 doi: 10.1093/nar/28.1.235
- 29.Eswar N, Webb B, Marti-Renom MA, Madhusudhan MS, Eramian D, Shen M-y, et al. (2006) Comparative protein structure modeling with Modeller. Published Online: John Wiley & Sons, Inc.
- 30.Pettersen EF, Goddard TD, Huang CC, Couch GS, Greenblatt DM, Meng EC, et al. (2004) UCSF chimera—A visualization system for exploratory research and analysis. J Comput Chem 25: 1605–1612. pmid:15264254 doi: 10.1002/jcc.20084
- 31.Bieth J, Spiess B, Wermuth CG (1974) Synthesis and analytical use of a highly sensitive and convenient substrate of elastase. Biochem Med 11: 350–357. pmid:4429553 doi: 10.1016/0006-2944(74)90134-3
- 32.Hummel BCW (1959) A modified spectrophotometric determination of chymotrypsin, trypsin, and thrombin. Can J Biochem Physiol 37: 1393–1399. pmid:14405350 doi: 10.1139/o59-157
- 33.Bergmeyer HU, Gawehn K, Grassl M (1974) Enzymes as biochemical reagents. New York: Academic Press.
- 34.Aramini JM, Huang YJ, Swapna GVT, Cort JR, Rajan PK, Xiao R, et al. (2007) Solution NMR structure of Escherichia coli ytfP expands the structural coverage of the UPFO131 protein domain family. Proteins 68: 789–795. pmid:17523190 doi: 10.1002/prot.21450
- 35.Vranken WF, Boucher W, Stevens TJ, Fogh RH, Pajon A, Llinas P, et al. (2005) The CCPN data model for NMR spectroscopy: Development of a software pipeline. Proteins 59: 687–696. pmid:15815974 doi: 10.1002/prot.20449
- 36.Ulrich EL, Akutsu H, Doreleijers JF, Harano Y, Ioannidis YE, Lin J, et al. (2008) BioMagResBank. Nucleic Acids Res 36: D402–D408. pmid:17984079 doi: 10.1093/nar/gkm957
- 37.Brunger AT, Adams PD, Clore GM, DeLano WL, Gros P, Grosse-Kunstleve RW, et al. (1998) Crystallography & NMR system: A new software suite for macromolecular structure determination. Acta Crystallogr D Biol Crystallogr 54: 905–921. pmid:9757107doi: 10.1107/s0907444998003254
- 38.Cornilescu G, Delaglio F, Bax A (1999) Protein backbone angle restraints from searching a database for chemical shift and sequence homology. J Biomol NMR 13: 289–302. pmid:10212987 doi: 10.1007/s10858-007-9166-6
- 39.Sharma D, Rajarathnam K (2000) C-13 NMR chemical shifts can predict disulfide bond formation. J Biomol NMR 18: 165–171. pmid:11101221
- 40.Bhattacharya A, Tejero R, Montelione GT (2007) Evaluating protein structures determined by structural genomics consortia. Proteins 66: 778–795. pmid:17186527doi: 10.1002/prot.21165
- 41.Luthy R, Bowie JU, Eisenberg D (1992) Assessment of protein models with 3-dimensional profiles. Nature 356: 83–85. pmid:1538787 doi: 10.1038/356083a0
- 42.Sippl MJ (1993) Recognition of errors in 3-dimensional structures of proteins. Proteins 17: 355–362. pmid:8108378 doi: 10.1002/prot.340170404
- 43.Laskowski RA, Macarthur MW, Moss DS, Thornton JM (1993) Procheck—a program to check the stereochemical quality of protein structures. J Appl Crystallogr 26: 283–291. doi: 10.1107/s0021889892009944
- 44.Lovell SC, Davis IW, Adrendall WB, de Bakker PIW, Word JM, Prisant MG, et al. (2003) Structure validation by C alpha geometry: phi,psi and C beta deviation. Proteins 50: 437–450. pmid:12557186 doi: 10.1002/prot.10286
- 45.Holm L, Rosenström P (2010) Dali server: conservation mapping in 3D. Nucleic Acids Res 38: W545–W549. doi: 10.1093/nar/gkq366. pmid:20457744
- 46.Hemmi H, Kumazaki T, Yoshizawa-Kumagaye K, Nishiuchi Y, Yoshida T, Ohkubo T, et al. (2005) Structural and functional study of an Anemonia elastase inhibitor, a “nonclassical” Kazal-type inhibitor from Anemonia sulcata. Biochemistry 44: 9626–9636. pmid:16008348 doi: 10.1021/bi0472806
- 47.Campos ITN, Souza TACB, Torquato RJS, De Marco R, Tanaka-Azevedo AM, Tanaka AS, et al. (2012) The Kazal-type inhibitors infestins 1 and 4 differ in specificity but are similar in three-dimensional structure. Acta Crystallogr D Biol Crystallogr 68: 695–702. doi: 10.1107/S0907444912009067. pmid:22683792
- 48.van de Locht A, Lamba D, Bauer M, Huber R, Friedrich T, Kroger B, et al. (1995) Two heads are better than one: crystal structure of the insect derived double domain Kazal inhibitor rhodniin in complex with thrombin. EMBO J 14: 5149–5157. pmid:7489704
- 49.Schlott B, Wöhnert J, Icke C, Hartmann M, Ramachandran R, Gührs K- H, et al. (2002) Interaction of Kazal-type inhibitor domains with serine proteinases: biochemical and structural studies. J Mol Biol 318: 533–546. pmid:12051857 doi: 10.1016/s0022-2836(02)00014-1
- 50.Qasim MA, Lu SM, Ding J, Bateman KS, James MNG, Anderson S, et al. (1999) Thermodynamic criterion for the conformation of P1 residues of substrates and of inhibitors in complexes with serine proteinases. Biochemistry 38: 7142–7150. pmid:10353824 doi: 10.1021/bi990265u
- 51.Cherney MM, Qasim MA, Maynes JT, Laskowski M, James MNG (2005) Structure of the subtilisin Carlsberg–OMTKY3 complex reveals two different ovomucoid conformations. Acta Crystallogr D Biol Crystallogr 61: 580–588. pmid:15858268 doi: 10.1107/s0907444905004889
- 52.Hemmi H, Kumazaki T, Yamazaki T, Kojima S, Yoshida T, Kyogoku Y, et al. (2003) Inhibitory specificity change of the ovomucoid third domain of the silver pheasant upon introduction of an engineered Cys14−Cys39 bond. Biochemistry 42: 2524–2534. pmid:12614146 doi: 10.1021/bi026727c
- 53.Crooks GE, Hon G, Chandonia JM, Brenner SE (2004) WebLogo: A sequence logo generator. Genome Res 14: 1188–1190. pmid:15173120 doi: 10.1101/gr.849004
- 54.Schneider TD, Stephens RM (1990) Sequence logos—a new way to display consensus sequences. Nucleic Acids Res 18: 6097–6100. pmid:2172928 doi: 10.1093/nar/18.20.6097
- 55.Santi L, Silva WOB, Pinto AFM, Schrank A, Vainstein MH (2010) Metarhizium anisopliae host-pathogen interaction: differential immunoproteomics reveals proteins involved in the infection process of arthropods. Fungal Biology 114: 312–319. doi: 10.1016/j.funbio.2010.01.006. pmid:20943140
- 56.Osbrink WLA, Williams KS, Connick WJ, Wright MS, Lax AR (2001) Virulence of bacteria associated with the Formosan subterranean termite (Isoptera: Rhinotermitidae) in New Orleans, LA. Environ Entomol 30: 443–448. doi: 10.1603/0046-225x-30.2.443
- 57.Da Silva P, Jouvensal L, Lamberty M, Bulet P, Caille A, Vovelle F (2003) Solution structure of termicin, an antimicrobial peptide from the termite Pseudacanthotermes spiniger. Protein Sci 12: 438–446. pmid:12592014 doi: 10.1110/ps.0228303
- 58.Khademi S, Guarino LA, Watanabe H, Tokuda G, Meyer EF (2002) Structure of an endoglucanase from termite, Nasutitermes takasagoensis. Acta Crystallogr D Biol Crystallogr 58: 653–659. pmid:11914490 doi: 10.1107/s0907444902002366
- 59.Jeng W-Y, Wang N-C, Lin M-H, Lin C-T, Liaw Y-C, Chang W-J, et al. (2011) Structural and functional analysis of three β-glucosidases from bacterium Clostridium cellulovorans, fungus Trichoderma reesei and termite Neotermes koshunensis. J Struct Biol 173: 46–56. doi: 10.1016/j.jsb.2010.07.008. pmid:20682343
- 60.Jeng WY, Wang NC, Lin CT, Chang WJ, Liu CI, Wang AHJ (2012) High-resolution structures of Neotermes koshunensis beta-glucosidase mutants provide insights into the catalytic mechanism and the synthesis of glucoconjugates. Acta Crystallogr D Biol Crystallogr 68: 829–838. doi: 10.1107/S0907444912013224. pmid:22751668
For further details log on website :
http://journals.plos.org/plosone/article?id=10.1371/journal.pone.0125376









No comments:
Post a Comment