Published Date
Author

For further details log on website:
http://journals.plos.org/plosone/article?id=10.1371/journal.pone.0120745
- Published: March 25, 2015
- http://dx.doi.org/10.1371/journal.pone.0120745
Author
Abstract
While hybridization of an invasive species with a native species is a common occurrence, hybridization between two invasive species is rare. Formosan subterranean termites (Coptotermes formosanus) and Asian subterranean termites (C. gestroi) are both ecologically successful and are the two most economically important termite pests in the world. Both species have spread throughout many areas of the world due to human activity; however, their distributions overlap in only three narrow areas because of distinct ecological requirements. In south Florida, where C. formosanus and C. gestroi are both invasive, the dispersal flight seasons of both species overlapped for the first time on record in 2013 and 2014. Pairings of heterospecific individuals were readily observed in the field and C. gestroi males preferentially engaged in mating behavior with C. formosanus females rather than females from their own species. In the laboratory, heterospecific and conspecific pairings had an equal colony establishment rate, but heterospecific incipient colonies had twice the growth rate of conspecific incipient colonies, suggesting a potential case of hybrid vigor. As all pre-zygotic barriers were lifted between the two species in the field, the apparent absence of post-zygotic barriers in the laboratory raises the possibility for introgressive hybridization in south Florida. While laboratory observations remain to be confirmed in the field, and the alate hybrid fertility is currently unknown, our results raise a tangible concern about the hybridization of two major destructive pest species. Such hybridization would likely be associated with a new economic impact.
Figures
Citation: Chouvenc T, Helmick EE, Su N-Y (2015) Hybridization of Two Major Termite Invaders as a Consequence of Human Activity. PLoS ONE 10(3): e0120745. doi:10.1371/journal.pone.0120745
Academic Editor: Alex Córdoba-Aguilar, Universidad Nacional Autonoma de Mexico, MEXICO
Received: November 25, 2014; Accepted: January 26, 2015; Published: March 25, 2015
Copyright: © 2015 Chouvenc et al. This is an open access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited
Data Availability: All relevant data are within the paper and its Supporting Information files.
Funding: The authors have no support or funding to report.
Competing interests: The authors have declared that no competing interests exist.
Introduction
Habitat alteration and human transportation have favored the spread of species with invasive capabilities as they may easily adapt to modified niches [1–3]. The establishment of invasive organisms in non-native areas can result in heterospecific interactions between invasive and native species, with potential for hybridization [4]. Introgression resulting from such hybridization can have important ecological and evolutionary consequences on native populations [5–8], and often are facilitated by human activity [4,9]. There is also mounting evidence that warming environments resulting from climate change can be an important factor contributing to such hybridization, either by altering the species distribution, or temporally shifting the mating season of species [10].
While the hybridization of non-native species with native species has been documented in a wide range of organisms [11,12], including plants [13], amphibians [14], fishes [15], mammals [16] and insects [17], few cases of hybridization involving two invasive species in non-native areas have been described. One such case is the hybridization of two invasive fire ant species (Solenopsis invicta × S. richteri) where a hybrid zone is now fully established in the Southern United States [18,19]. To a lesser extent, gene introgression from the Africanized honey bee to European honey bee populations (Apis mellifera subspecies) has become a problem for human activity in North and South America [20]. Hybrid introgressions among non-native organisms have only been described in a couple of social insects with socioeconomic impacts. Here, we describe a potentially new case of hybridization between two invaders in another social insect group with major economic importance.
Many subterranean termite (Isoptera: Rhinotermitidae) species are considered “urban pests” due to their tendency to attack man-made structures [21], and some are now invasive throughout the world, increasingly causing structural damage [22]. The Formosan subterranean termite (Coptotermes formosanus) and the Asian subterranean termite (C. gestroi) are the two most destructive structural pests in the world and are responsible for most of the $40 billion annual economic impact from termite damage [21]. As social insects, mature Coptotermescolonies can reach more than a million individuals [23] with caste polymorphism and polyethism [24], and have underground foraging galleries reaching up to 100 m, making detection and control difficult [25,26].
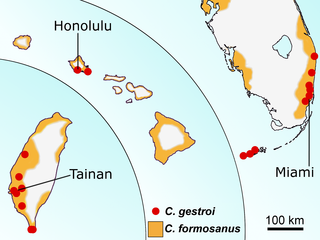
Coptotermes formosanus is endemic to China and Taiwan and has spread to many temperate and subtropical regions of the world [22]. It is now found throughout the southeastern United States and is responsible for more than $1 billion of structural damage each year in the United States alone [27]. Coptotermes gestroi is native to southeast Asia and has spread in many tropical regions, being potentially the most ubiquitous and destructive subterranean termite pest in the world [22]. Both species have distinct ecological requirements [28], but there are now established populations in many non-native areas due to human activity [29]. This observation reflects the current global biotic homogenization of some ecosystems, i.e. the replacement of native biotas by a small group of expanding non-native species in many parts of the world [30,31]. Their distributions now overlap in three narrow locations of the world [32,33]: the south part of the island of Taiwan, the island of Oahu in Hawaii, and southeast Florida (Fig. 1). However, studies concerning the interaction between C. formosanus and C. gestroi are restricted to competition between workers and soldiers from mature colonies, where individuals displayed interspecies agonism and competed for the access to resources [28,34]. The interspecies interactions of individuals from the reproductive caste (alates) have not yet been investigated.
Fig 1. The distributions of C. formosanus and C. gestroi overlap in three areas in the word.
From left to right: Taiwan, Hawaii, south Florida, according to [32,33] (modified for illustrative purpose only).
Swarming events (dispersal flights) in Coptotermes consist of mature colonies seasonally emitting thousands of alates at dusk [35]. Individuals drop their wings after the dispersal flight, find a mate, and engage in tandem behavior. The tandem behavior is initiated by the male as he maintains contact with the tip of the female’s abdomen [36]. The female then leads the way in search of a favorable nesting site, in which both individuals seclude themselves and establish the incipient colony [37]. It takes up to 8 yrs for these colonies to mature and initiate dispersal flights [38]. In south Florida, it was documented that the two species have distinct dispersal flight seasons [35]; however, for the first time on record, our monitoring of both species showed that alates swarmed simultaneously in a single location (Ft. Lauderdale, Florida, USA) on March 20th and April 3rd 2013.
Our preliminary observation of simultaneous swarming in 2013 implies that both geographical and temporal isolations may now be lifted between alates of C. formosanus and C. gestroi, giving the opportunity for interspecies mating. Because both species presumably went through allopatric divergence (different native distribution and ecological requirements), we hypothesized that the absence of reinforcement [39] may have prevented the formation of behavioral or physiological barriers against hybridization. We therefore investigated if pre-zygotic and post-zygotic barriers between C. formosanus and C. gestroi were lifted, and discussed the possibilities for the establishment of a hybrid termite population in areas where both species are established.
Results
(a) Swarming seasons overlap and tandem behavior
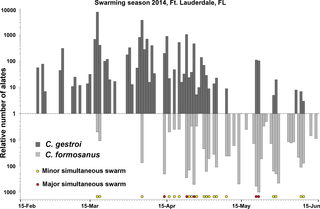
In 2013, simultaneous swarming events of C. formosanus and C. gestroi alates were casually observed on two distinct days (March 20th and April 3rd) at dusk in Ft. Lauderdale (Florida, USA). On those combined two days, 20–40 individuals from each sex were collected for both species and used for establishing incipient colonies in the laboratory. In 2014, the daily monitoring of swarming events at dusk, between February 15th and June 15th (120 d), resulted in observation of 55 swarming events for C. gestroi alates and 40 swarming events for C. formosanus alates at the same location. During this period, 20,018 C. gestroi alates and 5,001 C. formosanus alates were collected. There were 24 events of simultaneous dispersal flights between March 18th and June 9th, including five major simultaneous swarming events (Fig. 2), which confirmed the overlap of swarming seasons previously observed in 2013.
Fig 2. In south-east Florida, the dispersal flights (swarming seasons) of both species overlapped in 2014.
“Major simultaneous swarming event” indicates that more than 100 individuals were collected this day while “minor simultaneous swarming event” indicates that fewer than 100 individuals were collected for at least one of the species. Between March 18th and June 9th, there were 24 events of simultaneous C. formosanus and C. gestroi dispersal flights including 5 major simultaneous swarming events. The relative number of alates collected every day is presented on a log scale.
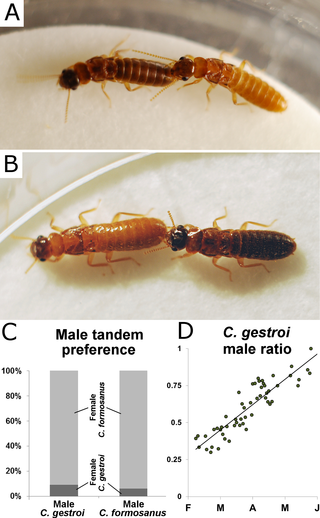
Males and females of both species readily initiated interspecies tandem behavior (Fig. 3A, 3B), with direct observations of interspecies tandems in the field (S1 Fig.). From a visual observation of termite tandems in the field on May 21st 2014, we estimated that interspecies tandems represented approximately one quarter of all tandem observations (38 individual interspecies tandems on a 3 m × 3 m surface within 30min of observation). However, we only observed male C. gestroi initiating and maintaining tandem with female C. formosanus in the field. In a laboratory choice assay between a female of each species, male C. gestroi and male C. formosanus both preferentially initiated a tandem with female C. formosanus (binomial test, 91% and 94% respectively, p<0.001 for both, Fig. 3C). In addition, toward the end of the C. gestroi swarming season, which corresponded to the peak of overlap of dispersal flights (April 15th—May 20th), C. gestroi sex-ratio was strongly male biased (Fig. 3D, linear model, F(42) = 133, p<0.001), increasing the chance for female C. formosanus and male C. gestroiinteractions in the field during simultaneous swarms.
Fig 3. Interspecies tandem.
(A) male C. formosanus maintaining tandem with female C. gestroi (B) male C. gestroimaintaining tandem with female C. formosanus (C) Tandem preference in a choice test, where male C. gestroi and male C. formosanus primarily initiated tandem behavior with female C. formosanus (binomial test, 91% and 94% respectively, p<0.001 for both). (D) Sex ratio was male-biased (linear model, F(42) = 133, p<0.001) for C. gestroi at the time of the swarming overlap, increasing the chance for male C. gestroi and female C. formosanus tandem formation in the field.
(b) Development of incipient hybrid colonies
Incipient colonies were established in 2013 in the laboratory by pairing a male and a female with four possible combinations, conspecific colonies (20 ♀C. gestroi × ♂C. gestroi, 18 ♀C. formosanus × ♂C. formosanus) and heterospecific colonies (10 ♀C. gestroi × ♂C. formosanus, 18 ♀C. formosanus × ♂C. gestroi) in individual rearing units. The initial colony development in all units was similar to what was previously described in C. formosanus incipient colonies [24]: the first eggs were observed within 25 d in most units and the first instar larvae developed within 40 d. First instar workers were observed within 65 d, and the first soldier was observed within 75 d (Fig. 4). Soldiers from mature colonies of C. gestroi and C. formosanus can be morphologically distinguished [40], however in incipient termite colonies, soldiers are produced from an accelerated developmental pathway (i.e. nanitic soldiers) and none of the soldiers displayed traits characteristic of either species [41].
Fig 4. Heterospecific colonies have viable hybrid offspring.
Shown here is a eight month-old incipient colony that contains the male C. gestroi, the female C. formosanus, eggs, larvae, workers and soldiers.
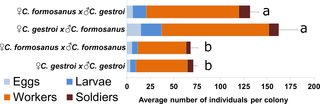
After one year of colony development, there was no significant difference in the failure of incipient colonies to establish between conspecific and heterospecific colonies (average colony mortality = 50%, typical under laboratory conditions [37], χ2 = 0.711, dl = 3, p = 0.87). However, one-year old heterospecific colonies displayed faster development (×2.03 the number of individuals produced in average) than conspecific colonies (F(3,32) = 24.21, p<0.001, Fig. 5). There was no difference in the proportion of castes and developmental stages among the four mating combinations after one year of development (χ2 test of independence, χ2 = 4.64, dl = 9, p = 0.86), with an average proportions of 6.8% eggs, 11.4% larvae, 74.1% workers and 7.7% soldiers.
Fig 5. After one year of development, heterospecific colonies have average 2.03 times the number of individuals than conspecific colonies (F(3,32) = 24.21, p<0.001, HSD post-hoc, same letter in a category indicates no significant difference).
(c) Molecular confirmation of the hybridization
Eight colonies (two per mating combination) were processed for genetic analysis. Each parental species displayed a single specific COII genotype, as previously described by [32]. For heterospecific colonies, the offspring inherited maternal mitochondrial markers, with ♀C. gestroi× ♂C. formosanus offspring displaying the C. gestroi COII genotype, and with ♀C. formosanus× ♂C. gestroi offspring displaying the C. formosanus COII genotype. Similarly, offspring from conspecific colonies displayed the COII type of their respective species (S1 Table).
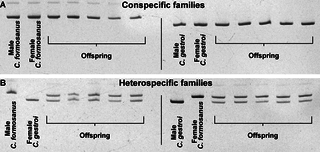
Microsatellite analysis for the Cg33 and Clac1 loci showed that all individuals from conspecific colonies were homozygous for both loci with species specific alleles (S2 Fig., S3 Fig.). We finally confirmed the hybridization of the offspring from heterospecific colonies (Fig. 6) as all individuals were heterozygous for Cg33 and Clac1 with alleles from both C. gestroi and C. formosanus origin (S2 Fig., S3 Fig.).
Fig 6. Results of the microsatellite Cg33 locus for the male, the female and five random offspring from the four mating combinations, where (A) conspecific colonies are homozygous for a species specific allele, and (B) heterospecific colonies have offspring displaying alleles from both species, confirming the hybridization.
Discussion
Our laboratory results indicate that C. formosanus and C. gestroi populations established in south Florida have the potential for the hybridization between two invasive species. The overlapping distribution and dispersal flight seasons in 2013 and 2014 provided the opportunity for male and female alates of the two species to interact directly. Interspecies mating was observed in the field and confirmed in the laboratory. All observations therefore indicate that there is currently no pre-zygotic barrier that may prevent hybridization between C. formosanusand C. gestroi in south Florida (S4 Fig.). The survival rate of hybrid colonies in the laboratory was not significantly different than that of conspecific colonies of the parental species. Although it was not possible to monitor the survival of incipient colonies in the field, our laboratory results suggest that heterospecific colonies have the potential for successfully establishing in the field. The genetic analysis of incipient colonies confirmed that individuals produced by heterospecific pairs were the result of hybridization, which excluded a potential case of parthenogenesis [42] or the unlikely storage of sperm by females from a previous mating [43]. Finally, hybrid colonies in the laboratory displayed high vigor by producing twice the number of individuals compared to incipient conspecific colonies.
At this point, it is premature to determine if any post-zygotic barriers would further prevent the hybridization process, but there are possible scenarios emerging from established heterospecific colonies. 1) Despite high colony vigor at the incipient stage, heterospecific colonies may later display maladaptive genotypes that would prevent them from reaching maturity or produce alates, eliminating all hybridization possibilities. 2) If produced, alates from heterospecific colony may be sterile, genetically or phenotypically incompatible with parental species, preventing any gene introgression back into the parental species. 3) If viable and fertile, alate hybrids may have limited overlapping swarming seasons with parental species, hindering the gene introgression process. 4) Finally, in the case of absence of post-zygotic barriers, with fully functional alate hybrids and their offspring, hybridization and introgression would be unavoidable.
Because male C. gestroi preferentially paired with female C. formosanus, we suggest that males of both species responded to the same cue for female attractiveness (presumably an identical sexual pheromone) but that female C. formosanus is a super releaser in comparison to female C. gestroi. In addition, the strong male-bias swarms of C. gestroi during the overlapping season also increased the chance for female C. formosanus and male C. gestroi mating combinations to found new colonies in the field. Such mating bias implies that in the scenario where all pre-zygotic and post-zygotic barriers were lifted, hybridization in the field would be unidirectional, resulting in an asymmetrical introgression of C. gestroi genes into C. formosanuspopulations.
There have been few studies regarding hybridization in termites and these have mostly described gene flow among populations of subspecies in the lower termites, Reticulitermes lucifugus [44] and Zootermopsis nevadensis [45], with cases of introgressive hybrids. In higher termites, no hybrid viability was observed between two species of Pseudacanthotermes [46], but some hybrid viability was observed in laboratory between two species of Nasutitermes for 60 d [47]. All cases with viable F1 occurred among termite populations with little genetic divergence (<30 substitutions in the COII gene). In comparison, Floridian populations of C. gestroi and C. formosanus have 78 substitutions for the COII gene [32] implying further genetic distant between both species than any previously observed hybridization in termites.
One of the paradoxes of invasive species is that they can be highly successful despite low levels of genetic diversity and social insects are good representatives of such a phenomenon [48]. Because both C. gestroi and C. formosanus were introduced in south Florida within the past 30 years [49], possibly with single introduction events, there were cycles of inbreeding that resulted in a relative absence of genetic diversity in such populations [50,51]. The two south Florida Coptotermes populations were therefore isolated from gene flow of their respective native genetic pool, and the accumulation of homozygous alleles may have resulted in sub-optimal vigor (inbreeding depression) [52]. It was documented that C. formosanus incipient colonies from their native range in China could reach more than 200 individuals within the first year [53], while in average we observed 70 individuals in our C. formosanus incipient colonies. The high numbers of individuals observed in hybrid colonies may reflect a case of heterosis (restoration of heterozygosity) [54], but such observation requires to be confirmed in the following years.
With a subtropical climate and strong human activity, the continuous spread of exotic termites in south Florida is inevitable [35]. Climate change can directly shift the distribution range [55] and the timing of reproduction of species [56] because of alterations of environmental conditions [57], and hybridization in animals has been observed as a consequence of shifts in species distributions due to climate change [10,58]. The unusually warm 2013 and 2014 winters (5thand 10th warmest winter on record) with successive cold fronts in the region (Source: NOAA) may have allowed for a wide overlap of the termite species dispersal flight seasons. In the scenario that such weather patterns become frequent in the near future [59], with the C. gestroirange predicted to move further north [60], simultaneous swarms may become a common occurrence. The documented case of introgressive hybridization in fire ants (Solenopsis) in the Southern United States [61] may serve as a cautionary tale, and the establishment of a hybridized Coptotermes population in south Florida with continuous gene flow between the two species remains a possibility. Both species have been established in Hawaii and Taiwan for a longer time [28,62], but hybridization has not been documented yet. The opportunity for C. formosanus and C. gestroi to hybridize may be unique to south Florida due to particular environmental cues and overlapping swarming seasons, but monitoring the dispersal flights in Hawaii and Taiwan would provide insight into the possibility of interspecies mating in all three locations.
To conclude, this study showed that all pre-zygotic barriers were lifted between C. formosanusand C. gestroi in the field, and high hybrid vigor was obtained in the laboratory. In the hypothetical scenario that no further post-zygotic barriers would prevent the hybridization, faster colony development in interspecific colonies (as observed in laboratory) would allow such colonies to reach maturity earlier and possibly disperse faster than the parental C. formosanusand C. gestroi colonies. Currently, it is not known if a hybrid colony can produce fertile and fully functional alates for maintaining hybrid populations and for possible introgressive hybridization back into the C. formosanus and C. gestroi populations (S4 Fig.). Because of the long life cycle of Coptotermes, it may take from 5 to 8 years before colonies mature in the laboratory and in the field [24,38], which would enable us to confirm the fertility of the F1 generation. In addition, due to the cryptic habit of termites and the currently unknown morphology of hybrid alates and soldiers from mature heterospecific colonies, it may take several years before the hybridization could be effectively monitored and confirmed in the field. Coptotermes mature colonies can contain millions of individuals and live up to 20 yrs [24] and even in the absence of alate hybrid fertility, the persistence of hybrid colonies in urban environments would still present a threat to structures (i.e., a kick from a mule is as good as a kick from a donkey).
Materials and Methods
(a) Termite collection and ethic statement
No specific permits were required for the termite’s collection from the field and for maintenance in laboratory. No endangered or protected species were involved in this study. All termites used for incipient colonies in this study for C. formosanus and C. gestroi were obtained from swarming events collected in Broward County (Florida, USA) in a privately owned, non-protected location.
In spring 2013, C. formosanus and C. gestroi alates were casually collected in Ft. Lauderdale, FL (26.105°N, 80.175°W) at dusk between March 15th and April 4th using a fluorescent light and an insect net. We obtained the first record of simultaneous C. formosanus and C. gestroiswarms during this time, as it was previously established that both species had distinct dispersal flight seasons [35]. However, we could only partially describe the flight phenology of the two species in the area for the 2013 swarm season. To fully characterize the flight activity of both species throughout the season, a light trap was setup daily between Feb 15th and June 15th 2014 at the same location. Monitoring was initiated 30 minutes before sunset and terminated 90 min after sunset. The light trap was modified from Peppuy [63] using a 1,720 lumen LED work light in addition to a fluorescent dark light and a regular fluorescent light. A 3m × 3m plastic sheet was laid under the light trap to ease observations around the trap. As termites were attracted to the light trap, we were able to observe tandem behavior from some individuals that landed outside the trap (S1 Fig.). Inside the trap, small rolls of moist corrugated cardboard were placed for temporary nesting sites, into which termites freely moved and isolated themselves in single cells. Cardboard rolls were transferred to the laboratory the next morning and processed to count the collected individuals, determine their sex and their species, and establish incipient colonies. In this study, we defined a “major simultaneous swarming event” as when both species swarmed on a given day with more than 100 individuals of both species collected in the light trap. A “Minor simultaneous swarming event” implies that less than 100 individuals were collected for at least one of the species.
(b) Tandem behavior assay
Tandem behavior is the process in which a male dealate follows a female dealate; the male antennae and mouthparts remain in contact with the female abdomen while she searches for a suitable nesting site [36,37]. Interspecies and intraspecies tandem behaviors were directly observed in the field on most evenings of simultaneous flights. In the laboratory, males were submitted to a choice test between a female of each species. One female C. gestroi and one female C. formosanus were placed in a 5 cm Petri dish with a moist filter paper on the bottom. One male C. gestroi or one male C. formosanus was introduced in the Petri dish and tandem behavior was monitored for 5 min (n = 100 males per species). If the male maintained a tandem position with a single female for at least 4 min (cumulative time), the female was marked as “preferred female.” (Note: as the males engaged in tandem behavior with a female of their choice, they maintained the tandem in 96% of the replicates. The eight remaining replicates were repeated to obtain a clear choice for all assays).
(c) Incipient colonies and rearing conditions
Dealates of both species were paired to establish incipient colonies in the laboratory. One male and one female were introduced into rearing units [24,64]. However, because of the relatively small number of individuals collected in 2013, a limited number of colonies were established per mating combination: 20 ♀C. gestroi × ♂C. gestroi, 18 ♀C. formosanus × ♂C. formosanus, 10 ♀C. gestroi × ♂C. formosanus, and 18 ♀C. formosanus × ♂C. gestroi. In 2014, a large number of alates were collected, and we established 200 incipient colonies per mating combination and will be used for future studies.
A rearing unit was composed of a transparent plastic cylindrical vial (8 cm × 2.5 cm diameter, internal volume = 37 cm3) with 6 g of moistened organic soil at the bottom. Four blocks of Picea sp. (5 cm × 0.5 cm × 0.5 cm) were positioned vertically and an additional Picea block (10 cm × 0.5 cm × 0.5 cm) was placed inside the vial, along the vertical side. A 3% agar solution was poured, leaving a 2-cm space at the top of the vial. When the agar was solidified, the long Picea block was removed from the vial to leave a hole in the agar, providing direct access to the soil on the bottom and to the wood. A perforated plastic cap was placed on the top to allow for aeration, but to limit desiccation and prevent escape. Rearing units were stored at 28°C for 365 d. A small amount of water was periodically added to vials that showed signs of dryness. After one year, all rearing units were opened to count for individuals of each caste and developmental stage (primary reproductives, eggs, larvae, workers and soldiers) and colonies that possessed a live male and female with their offspring were considered “successful colonies”.
(d) Sample preparation for DNA extractions
Two incipient colonies of each pairing combination (see “Incipient colonies and rearing conditions” above) were sacrificed for DNA extraction and subsequent polymerase chain reaction (PCR) for mitochondrial COII gene and microsatellites amplification. DNA was extracted from the parental male and female, 9 individual workers and one soldier from each of the incipient colonies using the Wizard DNA Extraction Kit (Promega, Inc.) per the manufacturer’s protocol, with the exception of the addition of GlycoBlue Coprecipitant (Invitrogen) for visualization of the DNA pellet. All DNA extraction concentrations were determined using a Qubit Fluorometer (Invitrogen) per the manufacturer’s protocol. DNA was stored at 4°C until ready for use.
(e) Mitochondrial COII PCR conditions
Mitochondrial COII gene amplification for 12 of the individuals extracted from each of the incipient colonies was completed using previously published mitochondrial COII primers [32]. Forward primer A-tLeu (5’-ATGGCAGATTAGTGCAATGG-3’) and reverse primer B-tLys (5’-GTTTAAGAGACCAGTACTTG-3’) were used to amplify C. formosanus; and forward primer C2F2 (5’-ATACCTCGACGWTATTCAGA-3) and reverse primer B-tLys were used to amplify C. gestroi, per reference [32] who indicated that using the A-tLeu/B-tLys primer combination for C. gestroi resulted in the amplification of a COII pseudogene.
Total COII PCR reactions, 50 μL, contained final reagent concentrations of 10X PCR buffer [65], 1.5 mM MgCl2, 200 mM each dNTP, 0.4μM each primer, 1.25 U Taq DNA polymerase (New England Biolabs, Inc., Ipswich, MA), and sterile ultrapure water, 100 ng of extracted DNA was used as template for amplification. PCR thermocycling was carried out using either Mastercycler Gradient Thermocycler (Eppendorf North America, Hauppauge, NY) or Arktik Thermocycler (Thermo Fisher Scientific, Inc., Waltham, MA) under the following conditions: initial denaturation at 95°C for 90 sec followed by 34 cycles of denaturation at 95°C for 60 seconds, annealing at 55°C for 60 sec, extension at 72°C for 2 min; and final extension at 72°C for 10 min. 10 μL of the resulting PCR products were electrophoresed on a 1% agarose gel, stained with ethidium bromide (EtBr) and visualized using UV illumination. The remaining 40 μL of PCR products were cleaned using a Wizard DNA Clean-Up System (Promega, Madison, WI, USA), quantified using Qubit Fluorometer (Invitrogen) and sent to the University of Florida’s Core Genomics Facilities (ICBR, Gainesville, FL) for sequencing.
Mitochondrial COII forward/reverse sequences were assembled and edited using DNA Baser v2.9 [66]. COII primers for C. gestroi amplified a fragment of 932 bp while primers for C. formosanus amplified a fragment of 765 bp. Consensus sequences were aligned using MEGA version 6 [67] and identified by BLAST analysis through NCBI website (http://blast.st-va.ncbi.nlm.nih.gov/Blast.cgi); resulting sequences for C. formosanus and C. gestroi matched accession numbers EU805757 and EU805770, respectively, which are both Florida accessions deposited in GenBank and described by [32]. Offspring for all combinations of incipient colonies carried their respective maternal COII sequences (S1 Table).
(f) Microsatellite genotyping
Twelve microsatellite markers previously described in various Coptotermes species [68–70] were investigated to determine whether any would be informative for cross-species amplification by testing them against a subset of individuals from the incipient colonies of C. formosanus and C. gestroi (S2 Table). Of the 12 microsatellite markers, Cg33 [68] and Clac1[70] were the only two loci that successfully yielded product where each colony displayed homozygous species specific alleles. These microsatellites were used for further analyses of all incipient colony mating combinations.
Total microsatellite PCR reactions, 50 μL, contained final reagent concentrations of 10X PCR buffer [64], 1.5 mM MgCl 2, 200 mM each dNTP, 0.4μM each primer, 1.25 U Taq DNA polymerase (New England Biolabs, Inc., Ipswich, MA), and sterile ultrapure water, 100 ng of DNA extractions were used as template for amplification. PCR thermocycling was carried out using either Mastercycler Gradient Thermocycler (Eppendorf North America, Hauppauge, NY) or Arktik Thermocycler (Thermo Fisher Scientific, Inc., Waltham, MA) under the following conditions for primers Cg33F/R and Clac1F/R: initial denaturation at 95°C for 90 sec followed by 34 cycles of denaturation at 95°C for 30 seconds, annealing at 56°C/53°C for 60 sec, extension at 72°C for 2 min; and final extension at 72°C for 8 min. Five μL of the resulting PCR products were electrophoresed on an 8% polyacrylamide gel (PAGE), stained with ethidium bromide (EtBr) and visualized using UV illumination. The individuals with strong amplification were then used in a follow-up PCR reaction under the same parameters as above for each of the primers but using fluorescent 6FAM-tagged forward primers. Again, PCR’s were checked for amplification using 5 μL of the resulting PCR products as described above. The remaining 45 μL of PCR products were either diluted 1:10 or 1:100 with ultrapure water and a 20 μL aliquot was sent to the University of Florida’s Core Genomics Facilities (ICBR, Gainesville, FL) for genotyping.
Geneious 7.1.5 (Biomatters Ltd.) was used to assemble microsatellite files and to score and bin alleles for each of the incipient colonies. The microsatellites were scored using the 3rd order least squares sizing method as recommended in the Geneious Microsatellite Plugin Manual (Biomatters Ltd.). Allele peaks with a Y-scale value of ≥8,000 were used in scoring alleles for all colonies; with the exception of some individuals where the initial PCR amplification itself was weak, these individuals alleles were scored if the Y-scale value was ≥4,000.
See S2 Fig. and S3 Fig. for microsatellite results. In brief, amplified locus Cg33 yielded two alleles at 192 bp for C. formosanus and 210 bp for C. gestroi; and Clac1 yielded 3 alleles at 173 or 183 bp in size for C. formosanus and 185 bp in size for C. gestroi. All offspring from the incipient colony pairings inherited one allele from each of the parental termites, confirming the hybrid status of the offspring.
(g) Statistical analysis
The tandem choice test between C. formosanus and C. gestroi females was analyzed with a binomial test for each male species (n = 100 per species). The variation of the male sex ratio in C. gestroi dispersal fights was analyzed with a linear model using R [71], using time as a factor and male sex ratio as a variable. The total number of individuals present in one-yr old colonies was compared between the four mating combinations using ANOVA (HSD post-hoc). Proportions of castes and developmental stages comparison between colonies of the four mating combinations was done using a χ2 test of independence.
Supporting Information

S1 Fig. Picture of tandem behavior between a female C. formosanus (left, large bright orange abdomen) and a male C. gestroi (right, small nutty brown abdomen) in the field.
Picture taken on May 21st 2014 at 8:32pm during a simultaneous swarm, 1.5 m away from the light trap used in the experiment. The two dealates were looking for a suitable nesting site while walking on a piece of spruce that was placed on top of the plastic tarp. Under poor lighting condition and moving objects, it was difficult to obtain sharp macro photography, but the clear difference of morphology allowed for immediate species identification. (Picture: T.C.).
doi:10.1371/journal.pone.0120745.s001
(TIF)
S2 Fig. Genotyping for the microsatellite locus Cg33 for 8 colonies (female, male, 9 workers and one soldier).
Hybrid offspring inherited specific alleles from their respective C. gestroi and C. formosanusparents.
doi:10.1371/journal.pone.0120745.s002
(TIF)
S3 Fig. Genotyping for the microsatellite locus Clac1 for 8 colonies (female, male, 9 workers and one soldier).
Hybrid offspring inherited specific alleles from their respective C. gestroi and C. formosanusparents.
doi:10.1371/journal.pone.0120745.s003
(TIF)
S4 Fig. Assessment for the risk of introgressive hybridization.
Our result supports that all pre-zygotic barriers are lifted, however the viability and fertility of F1alates needs to be confirmed. Monitoring for introgression back to parental populations is needed.
doi:10.1371/journal.pone.0120745.s004
(TIF)
S1 Table. COII genotypes of individuals from endogamous and exogamous colonies (2 replicates per mating combination).
Hybrid offspring inherited the maternal mitochondrial marker.
doi:10.1371/journal.pone.0120745.s005
(DOCX)
S2 Table. Microsatellites tested against C. gestroi and C. formosanus conspecific and heterospecific incipient colonies.
doi:10.1371/journal.pone.0120745.s006
(DOCX)
Acknowledgments
We thank Stephanie Osorio, Aaron Mullins, Lucas Carnohan, Sarah Bernard, Du He, Kelly Ugarelli and Ronald Pepin for technical assistance. We also thank Robin Giblin-Davis and Ed Vargo for insightful comments.
Author Contributions
Conceived and designed the experiments: TC EEH. Performed the experiments: TC EEH. Analyzed the data: TC. Contributed reagents/materials/analysis tools: NYS. Wrote the paper: TC EEH NYS.
References
- 1.Vitousek PM, D'Antonio CM, Loope LL, Rejmanek M, Westbrooks R. Introduced species: a significant component of human-caused global change. New Zealand J Ecol. 1997; 21: 1–16.
- 2.Suarez AV, Bolger DT, Case TJ. Effects of fragmentation and invasion on native ant communities in coastal southern California. Ecology. 1998; 79: 2041–2056. doi: 10.1890/0012-9658(1998)079[2041:eofaio]2.0.co;2
- 3.Leu M, Hanser SE, Knick ST. The human footprint in the west: a large-scale analysis of anthropogenic impacts. Ecol Appli. 2008; 18: 1119–1139. pmid:18686576 doi: 10.1890/07-0480.1
- 4.Crispo E, Moore JS, Lee-Yaw JA, Gray SM, Haller BC. Broken barriers: Human-induced changes to gene flow and introgression in animals. BioEssays. 2011; 33: 508–518. doi: 10.1002/bies.201000154. pmid:21523794
- 5.Huxel GR. Rapid displacement of native species by invasive species: effects of hybridization. Biol Conserv. 1999; 89: 143–152. doi: 10.1016/s0006-3207(98)00153-0
- 6.Mooney HA, Cleland EE. The evolutionary impact of invasive species. Proc Natl Acad Sci USA. 2001; 98: 5446–5451. pmid:11344292 doi: 10.1073/pnas.091093398
- 7.Ryan ME, Johnson JR, Fitzpatrick BM. Invasive hybrid tiger salamander genotypes impact native amphibians. Proc Natl Acad Sci USA. 2009;106: 11166–11171. doi: 10.1073/pnas.0902252106. pmid:19564601
- 8.Fitzpatrick BM, Johnson JR, Kump DK, Smith JJ, Voss SR, Shaffer HB. Rapid spread of invasive genes into a threatened native species. Proc Natl Acad Sci USA. 2010; 107: 3606–3610. doi: 10.1073/pnas.0911802107. pmid:20133596
- 9.Abbott R, Albach D, Ansell S, Arntzen JW, Baird SJE, Bierne N, et al. Hybridization and speciation. J Evol Biol. 2013; 26: 229–246. doi: 10.1111/j.1420-9101.2012.02599.x. pmid:23323997
- 10.Chunco AJ. Hybridization in a warmer world. Ecol Evol. 2014; 4: 2019–2031. doi: 10.1002/ece3.1052. pmid:24963394
- 11.Buggs RJA. Empirical study of hybrid zone movement. Heredity. 2007; 99: 301–312. pmid:17611495 doi: 10.1038/sj.hdy.6800997
- 12.Schierenbeck KA, Ellstrand NC. Hybridization and the evolution of invasiveness in plants and other organisms. Biol Invas. 2009; 11: 1093–1105. doi: 10.1007/s10530-008-9388-x
- 13.Paul J, Vachon N, Garroway C J, Freeland JR. Molecular data provide strong evidence of natural hybridization between native and introduced lineages of Phragmites australisin North America. Biol Invas. 2010; 12: 2967–2973. doi: 10.1007/s10530-010-9699-6
- 14.Riley SP, Bradley Shaffer H, Randal Voss S, Fitzpatrick BM. Hybridization between a rare, native tiger salamander (Ambystoma californiense) and its introduced congener. Ecol Appl. 2003; 13: 1263–1275. doi: 10.1890/02-5023
- 15.Winkler KA, Pammainger-Lahnsteiner B, Wanzenböck J, Weiss S. Hybridization and restricted gene flow between native and introduced stocks of Alpine whitefish (Coregonus sp.) across multiple environments. Mol Ecol. 2011; 20: 456–472. doi: 10.1111/j.1365-294X.2010.04961.x. pmid:21199024
- 16.Senn HV, Pemberton JM. Variable extent of hybridization between invasive sika (Cervus nippon) and native red deer (C. elaphus) in a small geographical area. Mol Ecol. 2009; 18: 862–876. doi: 10.1111/j.1365-294X.2008.04051.x. pmid:19175500
- 17.Kenis M, Auger-Rozenberg MA, Roques A, Timms L, Péré C, Cock MJ, et al. Ecological effects of invasive alien insects. In: Langor DW, Sweeney J, editors. Ecological impacts of non-native invertebrates and fungi on terrestrial ecosystems. Springer, Netherlands; 2009. pp. 21–45.
- 18.Vander Meer RK, Lofgren CS, Alvarez FM. Biochemical evidence for hybridization in fire ants. Florida Entomol. 1985; 68: 501–506. doi: 10.2307/3495147
- 19.Shoemaker DD, Ross KG, Arnold ML. Genetic structure and evolution of a fire ant hybrid zone. Evolution. 1996; 50: 1958–1976. doi: 10.2307/2410754
- 20.Scott Schneider S, DeGrandi-Hoffman G, Smith DR. The African honey bee: factors contributing to a successful biological invasion. Annu Rev Entomol. 2004; 49: 351–376. pmid:14651468 doi: 10.1146/annurev.ento.49.061802.123359
- 21.Rust MK, Su NY. Managing social insects of urban importance. Annu Rev Entomol. 2012; 57: 355–375. doi: 10.1146/annurev-ento-120710-100634. pmid:21942844
- 22.Evans TA, Forschler BT, Grace JK. Biology of invasive termites: a worldwide review. Annu Rev Entomol. 2013; 58: 455–474. doi: 10.1146/annurev-ento-120811-153554. pmid:23020620
- 23.Su NY, Scheffrahn RH. Foraging population and territory of the Formosan subterranean termite (Isoptera: Rhinotermitidae) in an urban environment. Sociobiology. 1988; 14: 353–360.
- 24.Chouvenc T, Su NY. Colony age-dependent pathway in caste development of Coptotermes formosanus Shiraki. Insect Soc. 2014; 61: 171–182. doi: 10.1007/s00040-014-0343-9
- 25.King EG, Spink WT. Foraging galleries of the Formosan subterranean termite, Coptotermes formosanus, in Louisiana. Ann Entomol Soc Am. 1969; 62: 536–542. doi: 10.1093/aesa/62.3.536
- 26.Su NY, Scheffrahn RH. A review of subterranean termite control practices and prospects for integrated pest management programmes. Integr Pest Manag Rev. 1998; 3: 1–13.
- 27.Corn ML, Johnson R. Invasive species: Major laws and the role of selected federal agencies. US Congressional Research Report. 2013; R43258.
- 28.Grace JK. Invasive termites revisited: Coptotermes gestroi meets Coptotermes formosanus. Proc.10th Pacific-rim termite research group conference. 2014; 1: 1–7.
- 29.Hochmair HH, Scheffrahn RH. Spatial association of marine dockage with land-borne infestations of invasive termites (Isoptera: Rhinotermitidae: Coptotermes) in urban south Florida. J Econ Entomol. 2010; 103: 1338–1346. pmid:20857745 doi: 10.1603/ec09428
- 30.Olden JD, LeRoy Poff N, Douglas MR, Douglas ME, Fausch KD. Ecological and evolutionary consequences of biotic homogenization. Trends Ecol Evol. 2004; 19: 18–24. pmid:16701221 doi: 10.1016/j.tree.2003.09.010
- 31.Su NY. How to become a successful invader. Florida Entomol. 2013; 96: 765–769. doi: 10.1653/024.096.0309
- 32.Li HF, Ye W, Su NY, Kanzaki N. Phylogeography of Coptotermes gestroi and Coptotermes formosanus (Isoptera: Rhinotermitidae) in Taiwan. Ann Entomol Soc Am. 2009; 102: 684–693. doi: 10.1603/008.102.0413
- 33.Woodrow RJ, Grace JK, Higa SY. Occurrence of Coptotermes vastator (lsoptera: Rhinotermitidae) on the Island of Oahu, Hawaii. Sociobiology. 2001; 38: 667–673.
- 34.Li HF, Yang RL, Su NY. Interspecific competition and territory defense mechanisms of Coptotermes formosanus and Coptotermes gestroi (Isoptera: Rhinotermitidae). Environ Entomol. 2010: 39: 1601–1607. doi: 10.1603/EN09262. pmid:22546458
- 35.Scheffrahn RH. Overview and current status of non-native termites (Isoptera) in Florida. Florida Entomol. 2013; 96: 781–788. doi: 10.1653/024.096.0311
- 36.Raina AK, Bland JM, Dickens JC, Park YI, Hollister B. Premating behavior of dealates of the Formosan subterranean termite and evidence for the presence of a contact sex pheromone. J Insect Behav. 2003; 16: 233–245.
- 37.Nutting WL. Flight and Colony foundation. In: Krishna K, Weesner FM, editors. Biology of termites Vol 1. Academic Press, New York; 1969. pp. 233–282.
- 38.Costa-Leonardo AM, Arab A, Hertel H. Note about the first swarming in a laboratory colony of Coptotermes gestroi (Isoptera: Rhinotermitidae). Sociobiology. 2005; 45: 137–139.
- 39.Servedio MR, Noor MA. The role of reinforcement in speciation: theory and data. Annu. Rev Ecol Evol Syst. 2003; 34: 339–364. doi: 10.1146/annurev.ecolsys.34.011802.132412
- 40.Scheffrahn RH, Su NY. Distribution of the termite genus Coptotermes (Isoptera: Rhinotermitidae) in Florida. Florida Entomol. 2005; 88: 201–203. doi: 10.1653/0015-4040(2005)088[0201:dottgc]2.0.co;2
- 41.Chouvenc T, Basile M, Su NY. Developmental instability in incipient colonies of social insects. 2014: PLoS ONE 9(11): e113949. doi: 10.1371/journal.pone.0113949. pmid:25423502
- 42.Matsuura K, Fujimoto M, Goka K. Sexual and asexual colony foundation and the mechanism of facultative parthenogenesis in the termite Reticulitermes speratus(Isoptera, Rhinotermitidae). Insect Soc. 2004; 51: 325–332. doi: 10.1007/s00040-004-0746-0
- 43.Raina A, Murphy C, Florane C, Williams K, Park YI, Ingber B. Structure of spermatheca, sperm dynamics, and associated bacteria in Formosan subterranean termite (Isoptera: Rhinotermitidae). Ann Entomol Soc Am. 2007; 100: 418–424. doi: 10.1603/0013-8746(2007)100[418:sossda]2.0.co;2
- 44.Lefebvre T, Châline N, Limousin D, Dupont S, Bagnères AG. From speciation to introgressive hybridization: the phylogeographic structure of an island subspecies of termite, Reticulitermes lucifugus corsicus. BMC Evol Biol. 2008; 8: 38. doi: 10.1186/1471-2148-8-38. pmid:18248672
- 45.Aldrich BT, Kambhampati S. Preliminary analysis of a hybrid zone between two subspecies of Zootermopsis nevadensis. Insect Soc. 2009; 56: 439–450. doi: 10.1007/s00040-009-0041-1
- 46.Connetable S, Robert A, Bordereau C. Dispersal flight and colony development in the fungus-growing termites Pseudacanthotermes spiniger and P. militaris. Insect Soc. 2012; 59: 269–277. doi: 10.1007/s00040-011-0216-4
- 47.Hartke TR, Rosengaus RB. Heterospecific pairing and hybridization between Nasutitermes corniger and N. ephratae. Naturwissenschaften. 2011; 98: 745–753. doi: 10.1007/s00114-011-0823-y. pmid:21761130
- 48.Tsutsui ND, Suarez AV, Holway DA, Case TJ. Reduced genetic variation and the success of an invasive species. Proc Natl Acad Sci USA. 2000; 97: 5948–5953. pmid:10811892 doi: 10.1073/pnas.100110397
- 49.Su NY, Scheffrahn RH, Weissling T. A new introduction of a subterranean termite, Coptotermes havilandi Holmgren (Isoptera: Rhinotermitidae) in Miami, Florida. Florida Entomol. 1997; 80: 408–411. doi: 10.2307/3495774
- 50.Yeap BK, Othman AS, Lee CY. Genetic analysis of population structure of Coptotermes gestroi (Isoptera: Rhinotermitidae) in native and introduced populations. Environ Entomol. 2011; 40: 470–476. doi: 10.1603/en10108
- 51.Husseneder C, Simms DM, Delatte JR, Wang C, Grace JK, Vargo EL. Genetic diversity and colony breeding structure in native and introduced ranges of the Formosan subterranean termite, Coptotermes formosanus. Biol Invas. 2012; 14: 419–437. doi: 10.1007/s10530-011-0087-7
- 52.Charlesworth D, Charlesworth B. Inbreeding depression and its evolutionary consequences. Annu Rev Ecol Syst. 1987; 18: 237–268. doi: 10.1146/annurev.es.18.110187.001321
- 53.Huang LW. Reproductive and physiological characteristics of the development of Coptotermes formosanus Shiraki colonies. Acta Entomologica Sinica. 1987; 30: 393–395.
- 54.Charlesworth D, Willis JH. The genetics of inbreeding depression. Nature Rev Genet. 2009; 10: 783–796. doi: 10.1038/nrg2664. pmid:19834483
- 55.Parmesan C, Yohe G. A globally coherent fingerprint of climate change impacts across natural systems. Nature. 2003; 421: 37–42. pmid:12511946 doi: 10.1038/nature01286
- 56.Primack RB, Higuchi H, Miller-Rushing AJ. The impact of climate change on cherry trees and other species in Japan. Biol Cons. 2009:142; 1943–1949. doi: 10.1016/j.biocon.2009.03.016
- 57.Walther GR, Post E, Convey P, Menzel A, Parmesan C, Beebee TJ, et al. 2002. Ecological responses to recent climate change. Nature. 2002; 416: 389–395. pmid:11919621 doi: 10.1038/416389a
- 58.Garroway CJ, Bowman J, Cascaden TJ, Holloway GL, Mahan CG, Malcolm JR, et al. Climate change induced hybridization in flying squirrels. Global Change Biol. 2010; 16: 113–121. doi: 10.1111/j.1365-2486.2009.01948.x
- 59.Coenen D. Projecting regional climate change in Florida via GIS-based downscaling of a general circulation model. Dissertation, University of Florida; 2010. 367p.
- 60.Li HF, Fujisaki I, Su NY. Predicting habitat suitability of Coptotermes gestroi (Isoptera: Rhinotermitidae) with species distribution models. J Econ Entomol. 2013; 106: 311–321. pmid:23448046 doi: 10.1603/ec12309
- 61.Ross KG, Shoemaker DD. Species delimitation in native South American fire ants. Mol Ecol. 2005; 14: 3419–3438. pmid:16156813 doi: 10.1111/j.1365-294x.2005.02661.x
- 62.Li HF, Su NY, Wu WJ. Solving the hundred-year controversy of Coptotermes taxonomy in Taiwan. Am Entomol. 2010; 56: 222–227. doi: 10.1093/ae/56.4.222
- 63.Peppuy A. Termites du nord Vietnam, communication chimique et isolement spécifique. Ph.D. dissertation, Université Pierre et Marie Curie, Paris VI. 1999; 146p.
- 64.Higa SY. Flight, colony foundation and development of the gonads of the primary reproductives of the Formosans subterranean termites, Coptotermes formosanusShiraki. Ph.D. dissertation. University of Hawaii, Honolulu. 1981; 173p.
- 65.Innis MA, Gelfand DH. Optimization of PCRs. In: Innis MA, Gelfand DH, Sninsky JJ, White TJ, editors. PCR Protocols: a Guide to Methods and Applications. San Diego, CA, USA. Academic Press. pp. 19903–12.
- 66.DNA Baser Sequence Assembler v2.9 Heracle BioSoft SRL. Available: http://www.dnabaser.com/ Accessed 2014 Nov 4.
- 67.Tamura K, Stecher G, Peterson D, Filipski A, Kumar S. MEGA6: molecular evolutionary genetics analysis version 6.0. Mol Biol Evol. 2013; 30: 2725–2729. doi: 10.1093/molbev/mst197. pmid:24132122
- 68.Yeap BK, Othman AS, Lee CY. Available: http://tomato.biol.trinity.edu/manuscripts/9-6/mer-09-0192.pdf. Accessed 2014 Jun 5.
- 69.Vargo EL, Henderson G. Identification of polymorphic microsatellite loci in the Formosan subterranean termite Coptotermes formosanus Shiraki. Mol Ecol. 2000; 9: 1935–1938. pmid:11091339 doi: 10.1046/j.1365-294x.2000.0090111935.x
- 70.Thompson GJ, Miller LR, Lenz M, Crozier RH. Phylogenetic analysis and trait evolution in Australian lineages of drywood termites (Isoptera, Kalotermitidae). Mol Phylogen Evol. 2000; 17: 419–429. pmid:11133196 doi: 10.1006/mpev.2000.0852
- 71.R Core Team. R Foundation for Statistical Computing, Vienna, Austria. Available: http://www.R-project.org/. Accessed 2015 Feb 9.
For further details log on website:
http://journals.plos.org/plosone/article?id=10.1371/journal.pone.0120745










No comments:
Post a Comment