Published Date
Author



For further details log on website :
http://journals.plos.org/plosone/article?id=10.1371/journal.pone.0104410
- Published: August 20, 2014
- http://dx.doi.org/10.1371/journal.pone.0104410
Author
Abstract
We describe here a co-occurrence (i.e. a syninclusion) of ants and termites in a piece of Mexican amber (Totolapa deposit, Chiapas), whose importance is two-fold. First, this finding suggests at least a middle Miocene antiquity for the modern, though poorly documented, relationship between Azteca ants and Nasutitermes termites. Second, the presence of a Neivamyrmex army ant documents an in situ raiding behaviour of the same age and within the same community, confirmed by the fact that the army ant is holding one of the termite worker between its mandibles and by the presence of a termite with bitten abdomen. In addition, we present how CT-scan imaging can be an efficient tool to describe the topology of resin flows within amber pieces, and to point out the different states of preservation of the embedded insects. This can help achieving a better understanding of taphonomical processes, and tests ethological and ecological hypotheses in such complex syninclusions.
Figures
Citation: Coty D, Aria C, Garrouste R, Wils P, Legendre F, Nel A (2014) The First Ant-Termite Syninclusion in Amber with CT-Scan Analysis of Taphonomy. PLoS ONE 9(8): e104410. doi:10.1371/journal.pone.0104410
Editor: Judith Korb, University of Freiburg, Germany
Received: February 21, 2014; Accepted: July 8, 2014; Published: August 20, 2014
Copyright: © 2014 Coty et al. This is an open-access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Funding: This study is supported by the French ISYEB, UMR 7205 CNRS, UPMC and EPHE, and the Spanish Ministry of Economy and Competitiveness project CGL2011-23948/BTE. The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript.
Competing interests: The authors have declared that no competing interests exist.
Introduction
Ants and termites represent ecologically critical organisms in intertropical and subtropical ecosystems, impacting by their abundance, organization and variety of occupied niches the availability of nutrients as well as the composition of soils [1]–[5]. Although conspicuous and ecologically meaningful, the relationships between these key eusocial insects are sparsely documented. The data gathered so far have reported on ant predatory behaviour over termites [6]–[9] and/or termite nest (termitaria) occupation by ants [10]–[13]. The question of the antiquity of these relationships remains untackled, which overlaps with elucidating the age and stability of modern ‘hot spots’ of biodiversity occupied by these insects. Ants and termites are recorded since the early Cretaceous [14], [15], but there is yet no fossil record of interactions between these two taxa. This is despite the fact that ants are very common in the Neogene Neotropical and Eocene Baltic amber [16]. They can be found in syninclusions with numerous other insects of various groups, therefore giving possibilities to track the origin of extant behaviours involving these organisms in the fossil record [17], [18]. The term ‘syninclusion’ is therefore intended here in the sens of Koteja [19], for multiple organic inclusions in the same piece of amber, essential for understanding arthropods paleobehaviours in past environments.
We aim here to describe the first syninclusion of termites and ants (for a review of syninclusions see [18]) in a piece of Mexican amber from the Totolapa deposit, together with an adult Psocodea (Figures 1A–B, S1, S2). The exceptional feature of this syninclusion lies in the fact that a raider ant (Neivamyrmex) and inquiline ants (Azteca) are entrapped together with Nasutitermes termites, thus ensuring that these genera were present at exactly the same time and shared at least a part of their ecological niche.
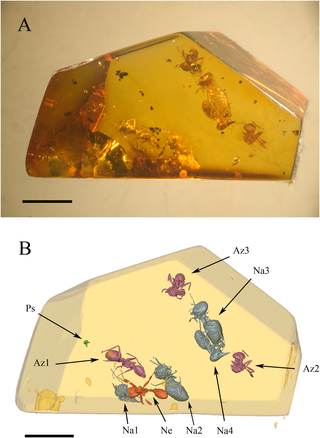
Figure 1. General configuration of the syninclusion.
(A) Overview of the amber piece, under optical microscope. scale bar = 3 mm. (B) Three-dimensional replica of the same; colours define taxonomic groups, viz. purple for Aztecaants, blue for Nasutitermes termites, red for Neivamyrmex ant, green for small Psocodea. Labels: Az1, Azteca ant nearest to predation scene; Az2 and Az3, two other Azteca ants, both trapped in a flow distinct to that of the others inclusions and whose physical density matches that of the Nasutitermes soldier; Na1, Nasutitermes worker trapped between the Neivamyrmex mandibles; Na2, isolated Nasutitermes termite closest to predation scene; Na3, Nasutitermes worker with damaged gaster; Na4, Nasutitermes soldier; Ne, Neivamyrmex ant; Ps, Psocodea; Scale bar = 3 mm.
Using CT-scan analysis with the purpose to improve access to taphonomically concealed features, we discovered that our amber piece was the result of several different flows and that the preservation of internal organic structures differed between insects. If tomographic analyses have already been widely used for taxonomical studies of insects, reconstructions of their external and internal morphology [20]–[26], and to illustrate a syninclusion in amber [27], we also use here the CT-scan as a tool to analyze the results of taphonomical processes in an amber syninclusion.
Material, Locality and Method
This amber piece was discovered in a batch of crude amber acquired by one of us (DC) from locals exploiting the Totolapa amber deposit (Salt River Mine). Later, the piece was offered to the Muséum National d’Histoire Naturelle de Paris (specimen MNHN.F.A49933). Totolapa is a village located in the central depression of Chiapas, 70 km south-east of Tuxtla Gutiérrez, the capital of Chiapas State. The Salt River amber mine, exploited since 2007 by Manuel Ramirez and his son Heriberto, is 1 km north of Totolapa, on the banks of the Salt River. The arthropod fauna collected by DC is currently under study. The age of the main Mexican amber locality, Simojovel, is still in debate, between Late Oligocene to Middle Miocene [28]–[33]. According to a geological map of the Instituto Nacional de Estatistica y Geografia [34] Totolapa amber would be Eocene in age, but a recent geological study of the Totolapa deposit suggests that the material originates from the Early Miocene Mazantic and Balumtum formations on top of Eocene marine facies [35]. As a matter of fact, Lambert et al. [36] suggested after a Carbon13 NMR spectroscopy study made on Baltic, Dominican and Mexican amber, that Simojovel and Totolapa amber came from the same palaeobotanical source; while Dominican amber, even if also closely related, shows more differences with both Mexican deposits. We therefore consider that the age of Totolapa amber is most probably between late Oligocene to middle Miocene and that the producing tree could also be Hymenaea mexicana [37] or a Hymenaea of undetermined species. Further geological studies are therefore needed to discover the precise age of Totolapa amber.
The original external surface of the amber piece has been removed by polishing; final lustration was done using diatomite powder. The specimens were examined under Nikon SZ10 and Olympus SZX9 stereomicroscopes. Photos were taken with an Olympus E-3 digital camera. Several digital pictures were reconstructed using Helicon Focus software.
X–ray tomography was realised in the AST-RX service (CT scan facility of the MNHN, UMS 2700), using a v|tome|x L240-180 from GE Sensing and Inspection Technologies phoenix|x-ray, with a X-Ray 180 KV/15 W nanofocus transmission tube, as well as a movable detector formed by a 20242 pixels (200 microns pixel). The voxel size of the reconstructed volume is 11.2 µm. 3D reconstructions and movies have been made using AVIZO 7.0 software. Variations in the material density in the amber piece are visible through changes of coloration from black (low density) to white (high density).
No permits were required for the described study.
Systematic palaeontology
Identifications of the specimens were possible at the generic level but not at the specific level, for the reasons indicated below.
Order Isoptera Brullé, 1832; Family Termitidae Latreille, 1802; Subfamily Nasutitermitinae Hare, 1937; Genus Nasutitermes Dudley, 1890; Nasutitermes species (Figures 1B, 2A–C).
Figure 2. Details of the syninclusion.
(A) General side view of the Neivamyrmex ant holding a Nasutitermes termite (Na1) between its mandibles, under optical microscope, scale bar = 1 mm. (B) detail of damaged gaster of Nasutitermes worker (Na3) closely contiguous to a Nasutitermessoldier (Na4), scale bar = 1 mm. (C) side view of closely contiguous Nasutitermes soldier (Na4) and worker (Na3), black arrow: digestive tube of Nasutitermes worker scale bar = 1 mm.
Comments. Typical workers and soldier termites assignable to Nasutitermes sp. by the following diagnostic characters: soldier with vestigial mandibles, with points; head capsule rounded, without constriction behind antennae; presence of a glabrous and narrow-tipped conical frontal tube (nasus); pronotum saddle-shaped and proctodeal segment not forming a loop on the right side of abdomen.
In Mexican amber, Nasutitermes was hitherto known only from imagos [38], to which our fossil soldier cannot be compared. From the two species known by soldiers in coeval Dominican amber, i.e. N. electronasutus Krishna, 1996 and N.rotundicephalus Krishna and Grimaldi, 1999, our fossil differs by its bare head as opposed to a head with long setae [39], [40]. Finally comparison of a new fossil Nasutitermes with the approximately 260 known modern species is highly difficult, given the absence of a reliable key and the uneven reliability of the various descriptions. We therefore do not ascribe our fossil to any particular species, and instead left it as Nasutitermes sp.
Order Hymenoptera Linné, 1758; Family Formicidae Latreille, 1809; Subfamily Dolichoderinae Forel, 1878; Genus Azteca Forel, 1878; Azteca species. (Figures 1A–B, 2A).
Comments. Dolichoderine ant with the following characters: nodiform petiole; unarmed hypostoma and propodeum; developed eyes; vertical first gastral tergite and anterior clypeal margin without a broad median concavity [41].
These Azteca ants will be described in a future paper, encompassing all the other Aztecapresent in the David Coty Totolapa amber collection.
Order Hymenoptera Linné, 1758; Family Formicidae Latreille, 1809; Subfamily Ecitoninae Forel, 1893; Genus Neivamyrmex Borgmeier, 1940; Neivamyrmex sp. (Figures 1A–B, 2A–C).
Comments. Ecitonine ant with the following diagnostic characters: eyes absent or reduced to an ommatidium; promesonotal suture absent or vestigial; antenna 12-segmented; antennal sockets fully exposed; absence of a preapical tooth on inner curvature of mid and hind pretarsal claws.
Only two fossil Ecitoninae, both from Dominican amber, are currently recorded: Neivamyrmex ectopus [42] and an undescribed army ant associated with a prey wasp pupa [43]. Neivamyrmex ectopus differs from our specimen in having a petiole with a subpetiolar process. Nevertheless, as the cuticle of our fossil specimen is badly preserved, and as we cannot reshape the amber piece (to preserve the syninclusion as a whole) in order to access further taxonomic details, we refrain from ascribing a new species.
General description of the amber piece
Our amber piece is 1.6 cm long, 1.0 cm wide and 1.2 cm high. It contains three Azteca ants (specimens Az1, Az2, and Az3 in Figure 1B), one Neivamyrmex ant (specimen Ne in Figure 1B), four Nasutitermes termites (Na1, Na2, Na3, and Na4, in Figure 1B), and a Psocoptera (specimen Ps in Figure 1B). The Neivamyrmex ant (Ne) holds a minor termite worker (Na1) between its mandibles (Figure 2A). In their vicinity, we can find one of the Nasutitermes workers (Na2) an Azteca ant (Az1), and a Psocoptera (Ps). Further away are grouped together two contiguous Nasutitermes termites, a soldier (Na4) with preserved digestive tube (Figure 2C), and a worker (Na3) with the gaster partly damaged (Figure 2B). The two remaining Azteca ants (Az2 and Az3) stand aside from both groups, and one from the other. See Figure S1 for a 3D view of the syninclusion.
CT-scan results
X-ray tomographic analysis revealed that our amber piece is in fact made of eight distinct layers corresponding to different flows, and that the distribution of the insects does not reflect a synchronous event. The layers are delimited by sinuose surfaces whose intersections with the different tomographic slicing are rendered as sinuate lines (variations of matter density visible in the images, see Figures 3, and S2).
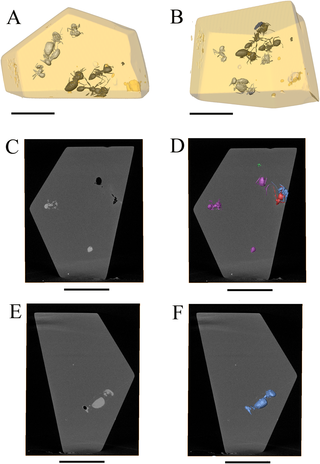
Figure 3. Virtual slicing from CT scan analysis showing flow boundaries.
Yellow arrows shows starting and ending points of flows limits. Na2 and Na3 belong to the same flow, strong disparity in density matter between specimens from same taxonomic groups, scale bar = 5 mm.
The fossil specimens are entrapped in two of the eight visible flows hereby identified (Figures 3, S2). Other flows are devoid of insect inclusions. The first flow with inclusions, herein named the ‘predation flow set’, contains the Neivamyrmex ant holding the minor termite between its mandibles (Ne+Na1), one Azteca ant (Az1), one Nasutitermes worker (Na2) the Psocoptera (Ps), and the two contiguous Nasutitermes termites (Na3+Na4). The second flow, named the ‘Azteca flow set’ contains the two isolated Azteca specimens (Az2 and Az3).
CT-scan analysis also emphasised variations in the physical density of the specimens. Empty specimens appear in black (low density registered) on the slices obtained with the CT-scan. In our case, the Neivamyrmex ant (Ne), the minor termite worker trapped between its mandibles (Na1), one Nasutitermes worker (Na2), and one Azteca ant (Az1) appear as empty structures inside the amber piece (in black on the slices, see Figures 3, 4C). On the contrary, the Nasutitermes soldier (Na4), the Nasutitermes worker with the damaged gaster (Na3) and two Azteca ants (Az2 and Az3) appear as full structure, denser than the amber (in white and light gray on the slices, see Figures 3, 4C–F). Thus, two coherent density sets can be distinguished in our piece of amber: the ‘Nasutitermes soldier density set’ and the ‘Neivamyrmex density set’ (Figures 4A–B).
Figure 4. Density differences identified by CT scan.
(A–B) Three-dimensional reconstruction artificially coloured representing two different density sets. ‘Nasutitermes soldier’ set in white, ‘Neivamyrmex’ set in black, as in CT-scan slices, Scale bar = 5 mm. (C) reslice of the original CT scan slice showing density differences between specimens. High density in white or light gray, low density in black or dark gray (darker than amber). Az2 and Az3 clearly denser than amber, Az1 and Ne empty structures, scale bar = 5 mm. (D) same slice as in B, but with 3D reconstruction of specimens, showing slice position in syninclusion, and specimen correspondences, scale bar = 5 mm. (E) reslice of original CT-scan slices, showing Nasutitermes soldier (Na4) and Nasutitermes worker (Na3), both denser than amber, scale bar = 5 mm. (F) same slice than E, but with 3D reconstruction of specimens, showing positions of specimens, scale bar = 5 mm.
Discussion
Ants-termites interactions – palaeoecological interpretation of the syninclusion
In the Neotropics, Nasutitermes is often involved in relationships with ants, possibly in relation with the fact that it is the termite genus with the highest number of species building conspicuous nests [4], [11]. A total of 54 extant ant species have been reported living in the different stages of the Nasutitermes nests [11]. Among those, Azteca species (A. chartifex Forel, 1896 and A. gnava Forel, 1906) have been found living at the three categorized termitaria stages (active, decadent, abandoned). The implications of extant Azteca in the opportunistic occupation of termitaria have been briefly described elsewhere [10], [11], but the nature of interactions is mostly unknown.
In the modern Venezuelan forests, it seems that some ants (including undetermined Aztecaspp.) occupy the nests of Nasutitermes corniger (Motschulsky, 1855) to protect themselves during the flooding events of the wet season [10]. During this temporary association, termites tolerate the predation of ants on their colony, as they themselves take the opportunity to feed on dead ants which constitute a valuable source of nitrogen. Nutrients flows occur both way between termites and ant in such association. Termites may also take benefits of the presence of inquilines ants in their nests to defend their common colony against predators [1], [44]–[46].
Cases of termites occupying parts of an active ant colony are also known [47]–[50]. Reasons of such cohabitations are generally unknown, although it has been noted that the contacts between ants and termites are rare in such cases, and qualified as neutral. Trager [50]mentioned that this type of association is frequent in the Neotropical region, involving different species of termites and ants.
Also called ‘The Huns of the insect world’ [51], all the modern species of army ants are major predators of both invertebrates and vertebrates [52]–[54]. They are also known to have a preference for preying on other eusocial insects, and ants in particular [49], [54]–[56]. Cases of predation of army ants on Nasutitermes are recorded in the Neotropics [57]. As mentioned by Brady [58], army ants ‘never hunt or forage solitarily’ but ‘dispatch a mass of cooperative, leaderless foragers to locate and overwhelm prey simultaneously’.
By phylogenetic inference, it is possible to say that our Mexican amber Neivamyrmex had the same behaviour and biology as its modern relatives (see [59], for inferences from recent fossil). The fact that this Neivamyrmex army ant holds a minor termite between its mandibles (Figure 2A) supports this hypothesis. Also, the gaster of the Na3 Nasutitermes termite is damaged, clearly showing traces of an ant bite (size and shape of the bite marks visible in Figure 2B). Lastly a phylogenetic inference shows that the Azteca ants were not predating the termites but more likely living with them in the same nest, as for their modern relatives. The presence of a Nasutitermes soldier (generally confined inside the nest and surging out of it for defence purpose), contiguous to the termite worker that exhibits an ant bite, also enhance the hypothesis (here again for an inference of a modern behaviour, see [59]) that the resin flowed close to a Nasutitermes nest.
This beam of evidences suggests that the fossil army ant present in our amber piece was part of a raid, during which the Nasutitermes termites and Azteca ants, sharing the same nest or interacting in some other degree might have been attacked, as it can typically occur in modern Neotropical settings.
The predation scene could also be the result of a peculiar type of scavenger behaviour, viz. when a predator is embedded while eating a dead insect only partly embedded in resin. However, the tomography shows that there is no discontinuity (a limit between two flows) between these two animals (see below for the study of flows using tomographic slices), which implies that they have been trapped in the same resin flow and thus invalidates this hypothesis.
The stress occurring during the embedment of the living ant together with a termite could have caused it to bite the termite randomly, but in this case, the presence of another termite partly eaten is not explained.
The presence of numerous Azteca ants (three specimens) and Nasutitermes termites (four specimens) in this small piece of amber, together with a Neivamyrmex ant reduces the possibility that Azteca ants and Nasutitermes termites have been randomly entrapped in the resin. It suggests that they were defending their common nest against an army ant ‘raid’ or interacting in some other degree while attacked. A ‘raid’ of army ants is always a strong perturbation for these eusocial communities, against which they have to defend, which bring many individuals out of their nest, therefore enhancing the probability to have many specimens of these different taxa entrapped together in a resin flow on the tree trunk.
Interpretation of CT-scan results
Structure of the amber.
The fact that amber pieces are almost always the result of several flowing event has been pointed out by various authors [60], [61]. Although the flow boundaries are partly visible at the surface of the amber piece (Figure 5), they are within the piece only detectable through greenish or brownish translucent surfaces, which appear under some particular view points, and light orientations (Figure 5). The CT-scan analysis therefore represents an informative enhancement allowing the clear mapping of the topology of each flow (Figure S2).
Figure 5. Flow margins visible under optic microscope.
Yellow arrows: ‘green veils’ representing limits between two flows, blue arrows: ‘sinuate line’ visible on surface of the amber piece, scale bar = 5 mm.
We discard the possibility that those sinuate lines could be cracks that may have occurred during the biostratinomy or the diagenesis of the amber, for two main reasons: 1) cracks never form smooth sinuate surfaces running across all the amber pieces; 2) they never follow the shape of inclusions but often damage them.
The surfaces of the flows appearing in CT-scan slices are always denser (i.e. lighter in the CT scan images) than the surrounding amber (also observed on other CT-scan slices of different amber pieces), suggesting that micro-particles, denser than the resin (not visible under optic microscope), deposited onto the amber surface before the arrival of the succeeding flow, or that a very thin layer of amber is harder at the surface contact between the two flows, as the resin was consolidated by polymerisation through sunlight and wind. The flow surfaces are nonetheless very weakly defined on the CT-scan slices and mostly not visible under optic microscope, possibly suggesting that the delays between flows might have been extremely short to limit the effects of polymerization on the flow surface, and avoid the deposition of dusts and debris on the fresh resin. It would therefore be likely that at least some relative degree of relation could exist between the density level of flow margins (in CT-scan imaging) and the time they were exposed to external elements, but further study on other material will be necessary to confirm such a hypothesis.
Differences of density between specimens.
Since the relative density of the insects is unrelated to their taxonomy (Azteca ants and Nasutitermes termites are present in different density sets), and since there is a clear spatial homogeneity between the two density groups (Figures 4A–B), variations of physical density between the specimens themselves is likely to express taphonomical disparities. This may be related to differences in the preservation of the cuticle and the inner organic structures [61], and differences between the insects before their entombment in the fresh resin, i.e. dead versus living animals, animals with or without filled digestive tubes, etc. As a matter of fact, insects in amber are frequently empty, since most of the internal organic content is anaerobically degraded, as a result of autolysis and the activity of endogenous bacteria [62], [63]. On the other hand, the preservation of internal structures (i.e. the digestive tube in the gaster of the Nasutitermes soldier, Figure 2C) may be due to a phenomenon of dehydration before the complete embedment in resin, as it has been shown that a pre-entombment dehydration of the insects inhibits the latter degradation process of their body inside the resin [64], [65].
Following this logic, it would therefore be possible that the specimens of the ‘Nasutitermesdensity set’ where embedded already dead, while the specimens of the ‘Neivamyrmex density set’ where embedded alive.
It has to be noted that the presence of a white aureole surrounding the body of inclusions, considered to be a foam of microscopic bubbles by Mierzejewski [66] and Weitschat and Wichard [67] can also help in some cases to reveal which insects where trapped alive or dead in the resin [61], as it is possibly the result of an early diagenesis reaction between fluids, produced by decay and decomposition of labile tissues, with sugar and terpenes in the resin. However, such foams are rare in Mexican and Dominican ambers. In the present case, no foam is visible around our specimens to help us in our analysis.
The two homogenous density groups are not distinguishable under optic microscope. Some preserved internal organs are visible, but the level of preservation of the cuticles is difficult to evaluate. One particularly odd case is the Nasutitermes with the damaged gaster (Na3) as this gaster is obviously empty of any kind of internal structures, while CT-scan images show that its whole body is denser than the amber. The fact that the damaged part of the gaster exhibits a bite mark caused by an ant, and is entirely covered by what seems to be the edge of an air bubble (Figure 2B) strongly suggests that this specimen was dead before entombment in fresh resin. In fact, regarding what we mentioned above, if the two closely contiguous Nasutitermes(Na3 and Na4) are likely to have been both dead before entombment in the resin, the similarity of their density levels may not have the same taphonomical origin, and remain to be elucidated for specimen Na3 (Nasutitermes).
Biostratinomic processes (the period between the moment when resin is exuded from the tree to the moment when it is buried in sediment) could also be responsible for these differences in matter density, as great differences can occur between flows in term of duration of the flowing event (viscosity), degree of humidity of the air, level and time of exposure to air and to UV, etc. Such variations can therefore create disparities in the taphonomic process between inclusions present in different flows.
If we therefore cross-compare the density distribution between specimens with the different flows in which they were embedded, our amber piece shows that the two density groups almost fit with the distribution of the specimens inside the flows, except for the closely contiguous Nasutitermes soldier and the Nasutitermes worker with a damaged gaster, as they share the same density level as the two Azteca ants that belong to a different flow. Regarding what has been mentioned above, this result can be due to differences between the insects themselves before entombment in the resin.
Conclusion
Our study provides evidence that some degree of relationship between Azteca ants and Nasutitermes termites might have already existed in Central America during the late Oligocene-middle Miocene period, together with the predation of army ants on other eusocial insects in the same community. However, the condition that led to the apparition of such interactions and their stability through time are still to be elucidated.
We also show here that beside anatomic reconstruction, CT-scan imaging can be used to study the taphonomy of syninclusions by allowing a more exhaustive description of resin flow topology and sequences, as well as a ‘cartography’ of density patterns of biotic inclusions. The main question to further address is to know in which measure both flow structures of the amber pieces and physical density variations of insect bodies can help to further reconstruct necrolysis, biostratinomic and diagenetic processes that occurred in the amber and its inclusions.
Supporting Information

Figure S2.
Movie of the syninclusion, showing the different flows of the resin that comprise the amber piece.
doi:10.1371/journal.pone.0104410.s002
(MPG)
Acknowledgments
We thank the two anonymous referees for their useful comments on the first version of the paper. DC thanks his friends, Manuel Ramirez and his son Heriberto, owners and miners of Salt River Mine, and their family, for their great help in his field research. We also thank Miguel Garcia Sanz (MNHN UMS 2700) for operating CT scan analysis.
Author Contributions
Conceived and designed the experiments: DC CA AN. Performed the experiments: DC RG PW AN. Analyzed the data: DC CA RG PW FL AN. Contributed reagents/materials/analysis tools: DC CA RG PW FL AN. Wrote the paper: DC CA RG PW FL AN.
References
- 1.Hölldobler B, Wilson EO (1990) The ants. Belknap Harvard University Press, Springer (publ.), Cambridge, Massachusetts, USA: [xiii] + 732.
- 2.Kaspari M (2001) Taxonomic level, trophic biology, and the regulation of local abundance. Global Ecology & Biogeography 10: 229–244. doi: 10.1046/j.1466-822x.2001.00214.x
- 3.Lavelle P, Bignell D, Lepage M (1997) Soil function in a changing world: the role of invertebrate ecosystem engineers, European Journal of Soil Biology. 33: 159–193.
- 4.Martius C (1994) Diversity and ecology of termites in Amazonian forests. Pedobiologia 38: 407–428.
- 5.Bignell DE, Roisin Y, Lo N (2010) Biology of termites: a modern synthesis. Springer publ: 576.
- 6.Deligne J, Quennedey A, Blum MS (1981) The enemies and defense mechanisms of termites. In Hermann, H. R. (ed.), Social Insects. Volume II, Academic Press, New York: 1–76.
- 7.Gonçalves TT, Ronaldo R Jr, DeSouza O, Ribeiro SP (2005) Predation and interference competition between ants (Hymenoptera: Formicidae) and arboreal termites (Isoptera: Termitidae). Sociobiology 46: 1–11.
- 8.Wilson EO (1971) The insect societies. Harvard University Press, Massachusetts: 562.
- 9.Cornelius ML, Grace JK (1995) Laboratory evaluations of interactions of three ant species with the Formosan subterranean termite (Isoptera: Rhinotermitidae). Sociobiology 26: 291–298.
- 10.Jaffe K, Ramos SC, Issa S (1995) Trophic interaction between ant and termites that share common nests. Annals of the Entomological Society of America 88: 328–333.
- 11.Santos PP, Vasconcellos A, Jahyny B, Delabie JHC (2010) Ant fauna (Hymenoptera, Formicidae) associated to arboreal nests of Nasutitermes spp. (Isoptera, Termitidae) in a cacao plantation in southeastern Bahia. Revista Brasileira de Entomologia 54: 450–454. doi: 10.1590/s0085-56262010000300016
- 12.Leponce M, Roisin Y, Pasteels JM (1999) Community interactions betweens ants and arboreal-nesting termites in New Guinea coconut plantations. Insectes Sociaux 46: 126–130. doi: 10.1007/s000400050122
- 13.Quinet Y, Tekule N, De Biseau JC (2004) Behavioural interactions between Crematogaster brevispinosa rochai Forel (Hymenoptera: Formicidae) and two Nasutitermes species (Isoptera: Termitidae). Journal of Insect Behavior 18: 1–17.
- 14.Perrichot V, Lacau S, Néraudeau D, Nel A (2008) Fossil evidence for the early ant evolution. Naturwissenschaften 95: 85–90.
- 15.Krishna K, Engel MS, Grimaldi DA, Krishna V (2013) Treatise on the Isoptera of the world. Bulletin of American Museum of Natural History 377: 2704. doi: 10.1206/377.6
- 16.LaPolla JS, Dlussky GM, Perrichot V (2013) Ants and the fossil record. Annual Review of Entomology 58: 609–630. doi: 10.1146/annurev-ento-120710-100600
- 17.Perkovsky EE, Rasnitsyn AP, Vlaskin AP, Rasnitsyn SP (2012) Contribution to the study of the structure of amber forst communities based on analysis of syninclusions in the Rovno amber (Late Eocene of Ukraine). Paleontological Journal 46: 293–301. doi: 10.1134/s0031030112030136
- 18.Boucot AJ, Poinar GO Jr (2010) Fossil behavior compendium. CRC Press, Taylor & Francis group, Boca Raton, FL, USA: 391.
- 19.Koteja J (1989) Syninclusions. Wrostek 8: 7–8.
- 20.Lak M, Néraudeau D, Nel A, Cloetens P, Perrichot V, et al. (2008) Phase contrast X-Ray synchrotron imaging: opening access to fossil inclusions in opaque amber. Microscopy and Microanalysis 14: 251–259. doi: 10.1017/s1431927608080264
- 21.Grimaldi D, Nguyen T, Ketcham R (2000) Ultra-high-resolution X-ray computed tomography (UHR CT) and the study of fossils in amber. 77–92, in D. Grimaldi (ed.), 2000.
- 22.Henderickx H, Cnudde V, Masschaele B, Dierick M, Vlassenbroeck J, et al. (2006) Description of a new fossil Pseudogarypus (Pseudoscorpiones: Pseudogarypidae) with the use of X-ray micro CT to penetrate opaque amber. Zootaxa 1305: 41–50.
- 23.Tafforeau P, Boistel R, Boller E, Bravin A, Brunet M, et al. (2006) Applications of X-ray synchrotron microtomography for non-destructive 3D studies of paleontological specimens. Applied Physics A: Materials Science & Processing 83: 195–202. doi: 10.1007/s00339-006-3507-2
- 24.Penney D, Dierick M, Cnudde V, Masschaele B, Vlassenbroeck J, et al. (2007) First fossil Micropholcommatidae (Araneae), imaged in Eocene Paris amber using X-ray computed tomography. Zootaxa 1623: 47–53.
- 25.Soriano C, Archer M, Azar D, Creaser P, Delclos X, et al. (2010) Synchrotron X-ray imaging of inclusions in amber. Comptes Rendus Palevol 9: 361–368. doi: 10.1016/j.crpv.2010.07.014
- 26.Sutton MD, Rahman I, Garwood R (2014) Techniques for virtual paleontology (analytical methods in Earth and environmental science). Wiley-Blackwell/John Wiley &Sons: i–viii + 1–200.
- 27.Penney D, McNeil A, Green DI, Bradley R, Jepson JE, et al. (2012) Ancient Ephemeroptera-Collembola symbiosis predicts contemporary phoretic associations. PLoS ONE 7(10): e47651.
- 28.Frost SH, Langenheim RL (1974) Cenozoic reef biofacies; Tertiary larger foraminifera and scleractinian corals from Chiapas, Mexico. Northern Illinois University Press, De Kalb, 1–388.
- 29.Ferrusquía-Villafranca I (2006) The first Paleogene mammal record of Middle America: Simojovelhyus pocitosense (Helohyidae, Artiodactyla). Journal of Vertebrate Paleontology 26: 989–1001. doi: 10.1671/0272-4634(2006)26[989:tfpmro]2.0.co;2
- 30.Castaneda-Posadas C, Cevallos-Ferriz SRS (2007) Swietenia (Meliaceae) flower in Late Oligocene–Early Miocene amber from Simojovel De Allende, Chiapas, Mexico. American Journal of Botany 94: 1821–1827. doi: 10.3732/ajb.94.11.1821
- 31.Solòrzano-Kraemer MM (2007) Systematic, palaeoecology, and palaeobiogeography of the insect fauna from Mexican amber. Palaeontographica (A) 282: 1–133.
- 32.Vega FJT, Nyborg T, Coutino MA, Solé JM, Hernández-Monzón O (2009) Neogene Crustacea from southeastern Mexico. Bulletin of the Mizunami Fossil Museum 35: 51–69.
- 33.Perrilliat MC, Vega FJ, Coutino MA (2010) Miocene mollusks from the Simojovel area in Chiapas, southwestern Mexico. Journal of the South American Earth Sciences 30: 111–119.
- 34.INEGI (1985) Carta Geológica, E15–11 (Tuxtla Gutiérrez), escala 1∶250,000. SPP/INEGI, Instituto Nacional de Estadística, Geografía y Informática, Mexico City, Mexico.
- 35.Durán-Ruiz C, Riquelme F, Coutiño-José M, Carbot-Chanona G, Castaño-Meneses G, et al. (2013) Ants from the Miocene Totolapa amber (Chiapas, México), with the first record of the genus Forelius (Hymenoptera, Formicidae). Canadian Journal of Earth Sciences 50: 495–502. doi: 10.1139/cjes-2012-0166
- 36.Lambert JB, Frye JS, Lee TA Jr, Welch CJ, Poinar GO Jr (1989) Analysis of Mexican amber by Carbon-13 NMR Spectroscopy. Archeological Chemistry 4: 381–388. doi: 10.1021/ba-1988-0220.ch021
- 37.Poinar GO Jr, Brown AE (2002) Hymenea mexicana spp. nov. (Leguminosae: Caesalpinioideae) from Mexican amber indicates Old World connections. Botanical Journal of the Linnean Society 139: 125–132.
- 38.Krishna K, Engel MS, Grimaldi DA, Krishna V (2013) Treatise on the Isoptera of the world. Bulletin of the American Museum of Natural History 377: 1–2704. doi: 10.1206/377.6
- 39.Krishna K (1996) New fossil species of termites of the subfamily Nasutitermitinae from Dominican and Mexican amber. American Museum Novitates 3176: 1–13.
- 40.Krishna K, Grimaldi DA (2009) Diverse Rhinotermitidae and Termitidae (Isoptera) in Dominican amber. American Museum Novitates 3640: 1–48.
- 41.Bolton B (1994) Identification guide to the ant genera of the World. Harvard University Press, Cambridge, London, Massachusetts, England: [iv] + 1–222.
- 42.Wilson EO (1985) Ants of the Dominican amber (Hymenoptera: Formicidae) 2. The first fossil army ants. Psyche 92: 11–16.
- 43.Poinar GO Jr, Poinar R (1999) The amber forest. A reconstruction of a vanished World. Princeton University Press, Princeton, New Jersey: i–xiii + 1–239.
- 44.Howse P (1984) Alarm defense and chemical ecology of social insect. In: Lewis, T. (ed.). Insect communication. Academic Press, London: 151–164.
- 45.Jolivet P (1986) Les fourmis et les plantes, un exemple de coévolution. Boubée Publ., Paris. 255.
- 46.Highashi S, Ito F (1989) Defense of termitaria by termitophilous ants. Oecologia 80: 145–147. doi: 10.1007/bf00380142
- 47.Wheeler WM (1936) Ecological relations of Ponerinae and other ants to termites. Proceedings of the National Academy of Sciences 71: 159–243. doi: 10.2307/20023221
- 48.Gray B (1974) Associated fauna found in a nests of Myrmecia (Hymenoptera, Formicidae). Insectes Sociaux 21: 289–300. doi: 10.1007/bf02226920
- 49.Sennepin A (1999) Symbioses entre fourmis et termites: structures et implications. Actes des Colloques des Insectes Sociaux 12: 181–190.
- 50.Trager JC (1991) A revision of the fire ants Solenopsis geminata group (Hymenoptera, Formicidae: Myrmicinae). Journal of the New York Entomological Society 99: 141–198.
- 51.Wheeler WM (1910) Ants, their structure, development and behavior. Columbia University Biological Series, New York 9: xxvi + 663.
- 52.Rettenmeyer CW (1963) Behavioral study of army ant. University of Kansas Science Bulletin 44: 281–465.
- 53.Schneirla TC (1971) Army ants: a study in social organization. W.H. Freeman & Co Ltd, San Francisco: 349.
- 54.Gotwald WH Jr (1995) Army ants: the biology of social predation. Cornell University Press, Ithaca, New York: 302.
- 55.LaPolla JS, Mueller UG, Seid M, Cover SP (2002) Predation by the army ant Neivamyrmex rugulosus on the fungus-growing ant Trachymyrmex arizonensis. Insectes Sociaux 49: 251–256. doi: 10.1007/s00040-002-8310-2
- 56.Le Breton J, Dejean A, Snelling G, Orivel J (2007) Specialized predation on Wasmannia auropunctata by the army ant species Neivamyrmex compressinodis. Journal of Applied Entomology 131: 740–743. doi: 10.1111/j.1439-0418.2007.01221.x
- 57.Souza JLP, Moura CAR (2008) Predation of ants and termites by army ants, Nomamyrmex esenbeckii (Formicidae, Ecitoninae) in the Brazilian Amazon. Sociobiology 52: 399–402.
- 58.Brady SG (2003) Evolution of the army ant syndrome: long-term evolutionary stasis of a complex of behavioral and reproductive adaptations. Proceedings of the National Academy of Sciences 100: 6575–6579. doi: 10.1073/pnas.1137809100
- 59.Nel A (1997) The probabilistic inference of unknown data in phylogenetic analysis. In: Grandcolas P (ed.). The origin of biodiversity in insects: phylogenetic tests of evolutionary scenarios. Mémoires du Muséum National d’Histoire Naturelle, Paris 173: 305–327.
- 60.Grimaldi D, Engel MS (2005) Evolution of the Insects. Cambridge University Press, New York/Cambridge, USA, 755 p.
- 61.Martínez-Delclòs X, Briggs DEG, Peñalver E (2004) Taphonomy of insects in carbonates and amber. Palaeogeography, Palaeoclimatology, Palaeoecology 3225: 1–46. doi: 10.1016/s0031-0182(03)00643-6
- 62.Allison PA, Briggs DEG (1991a) The taphonomy of softbodied animals. In: Donovan S.K. (Ed.), The Processes of Fossilization. Belhaven Press, London: 120–140.
- 63.Allison PA, Briggs DEG (1991b) Taphonomy of non-mineralized tissues. In: Allison PA, Briggs DEG, (Eds.), Taphonomy: Releasing the data locked in the fossil record. Plenum Press, New York: 25–70.
- 64.Henwood A (1992a) Insect taphonomy from Tertiary Amber of the Dominican Republic. Ph.D. Thesis, University of Cambridge 166.
- 65.Henwood A (1992b) Exceptional preservation of dipteran flight muscle and the taphonomy of insects in amber. Palaios 7: 203–212.
- 66.Mierzejewski P (1978) Electron microscopy study on the milky impurities covering arthropod inclusions in the Baltic amber. Prace Muzeum Zeimi 28: 81–84.
- 67.Weitschat W, Wichard W (1998) Atlas der Pflanzen und Tiere im Baltischen Bernstein. Pfeil, Munchen: 256.
For further details log on website :
http://journals.plos.org/plosone/article?id=10.1371/journal.pone.0104410









No comments:
Post a Comment